

Genome-wide association between YAP/TAZ/TEAD and AP-1 at enhancers drives oncogenic growth
- PMID: 26258633
- PMCID: PMC6186417
- DOI: 10.1038/ncb3216
Genome-wide association between YAP/TAZ/TEAD and AP-1 at enhancers drives oncogenic growth
Abstract
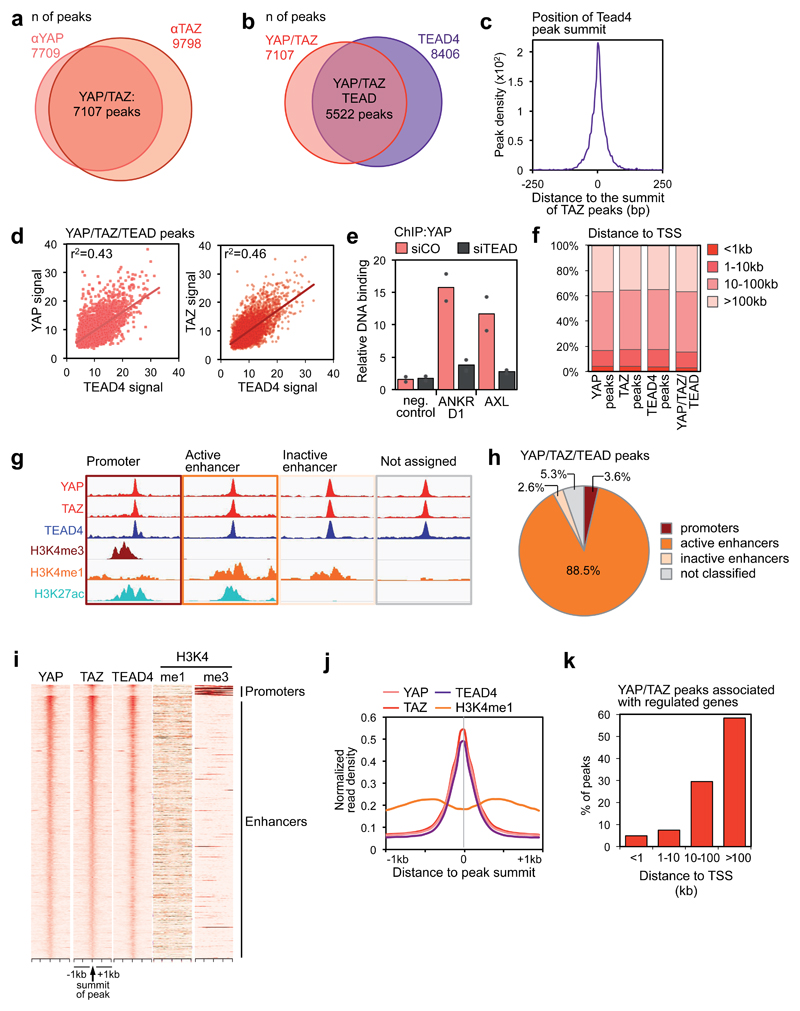
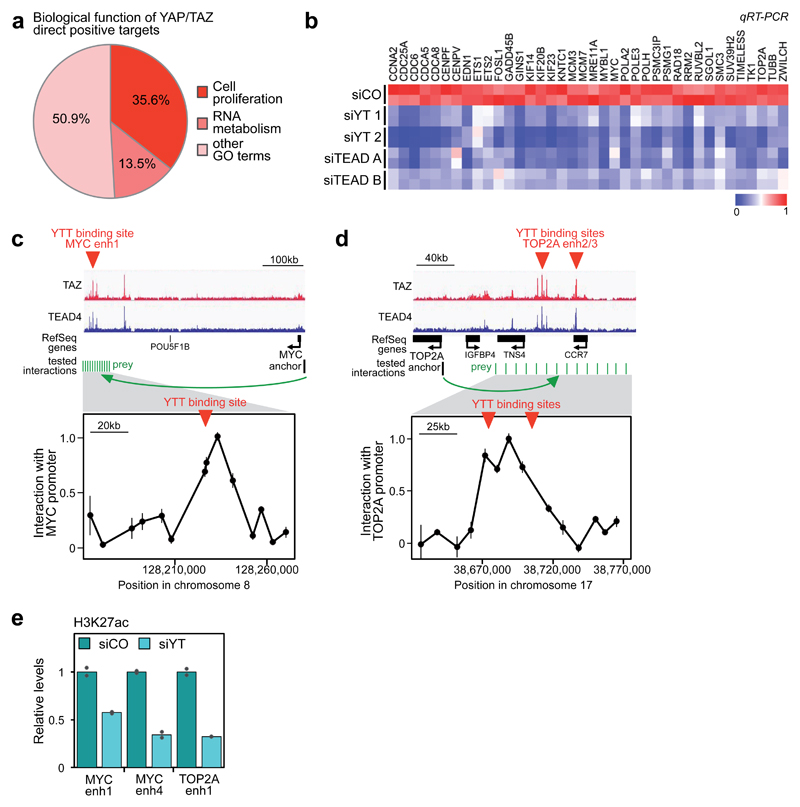
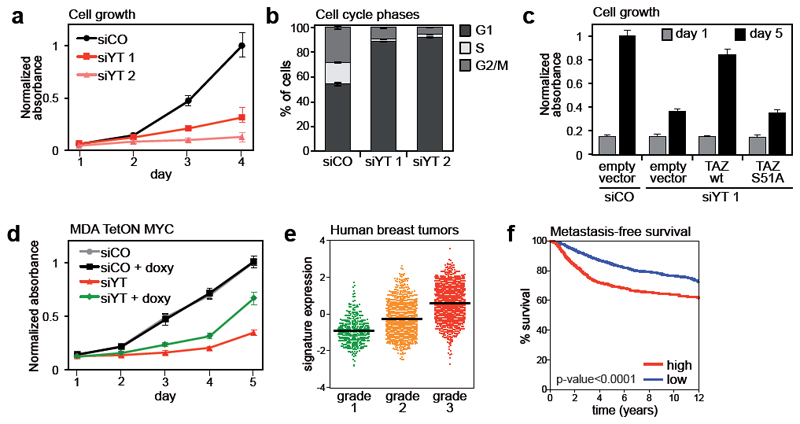
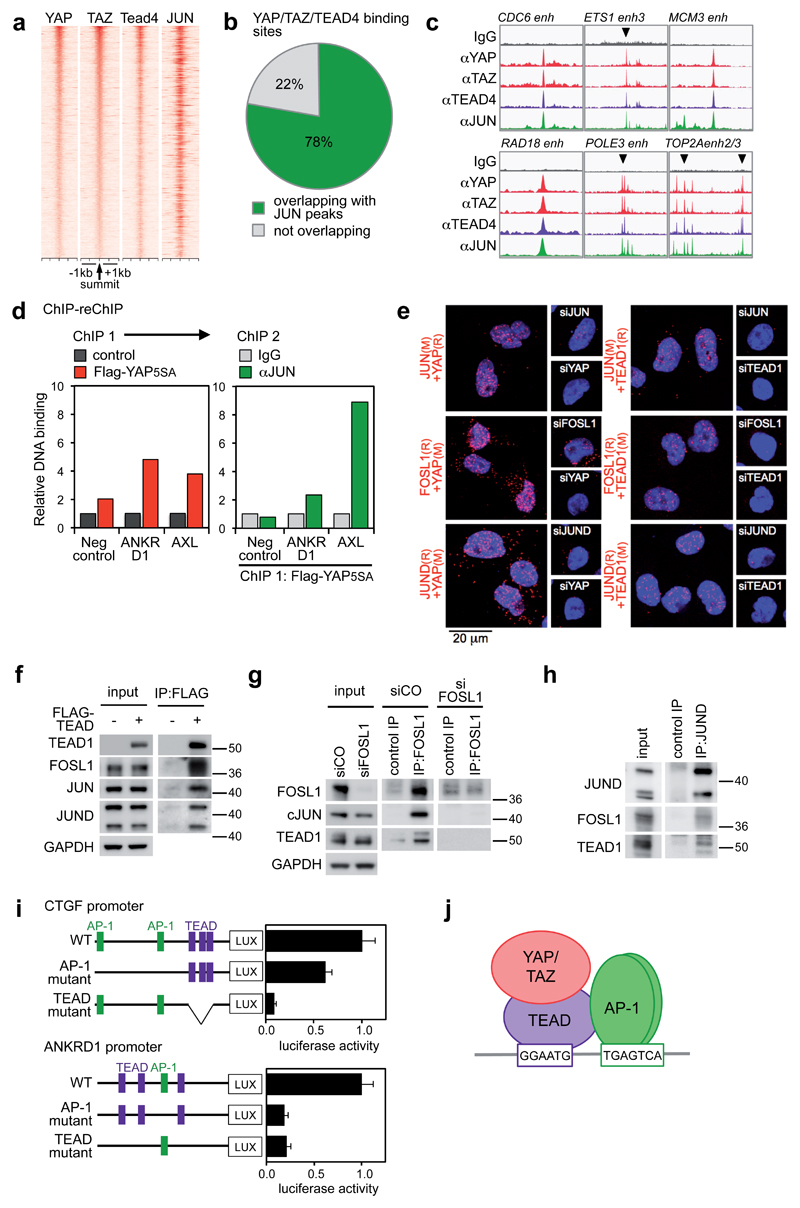
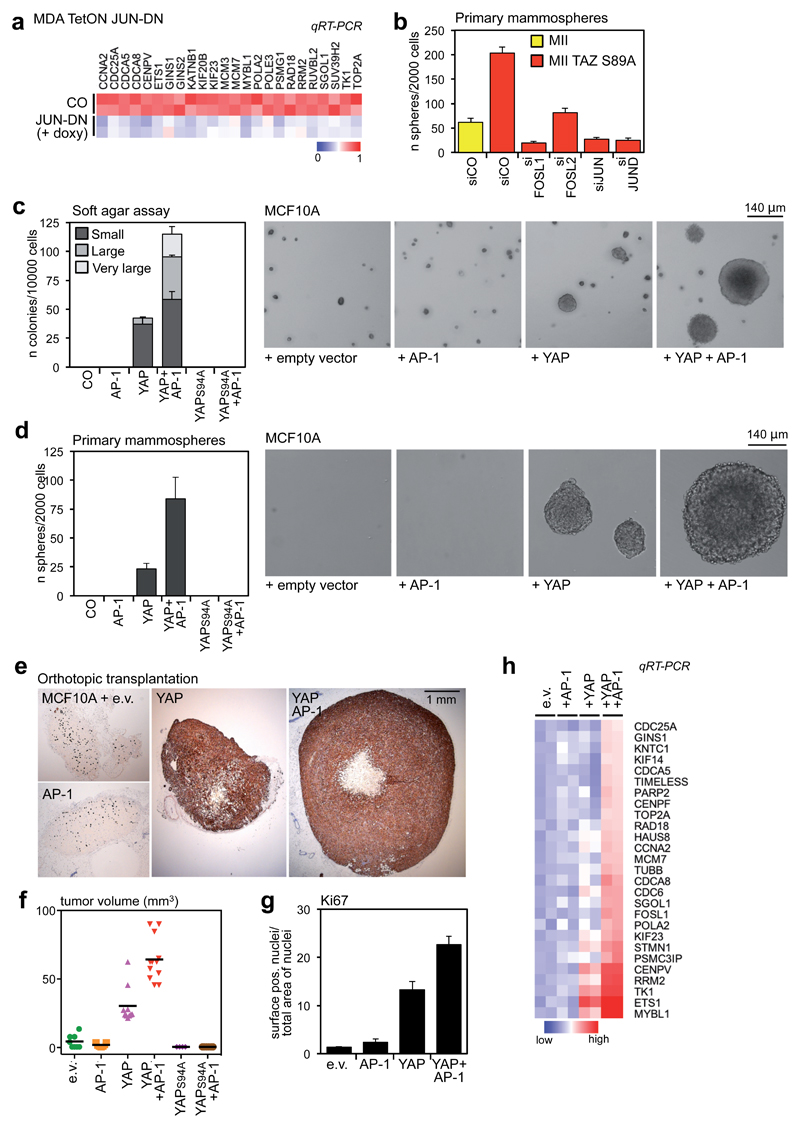
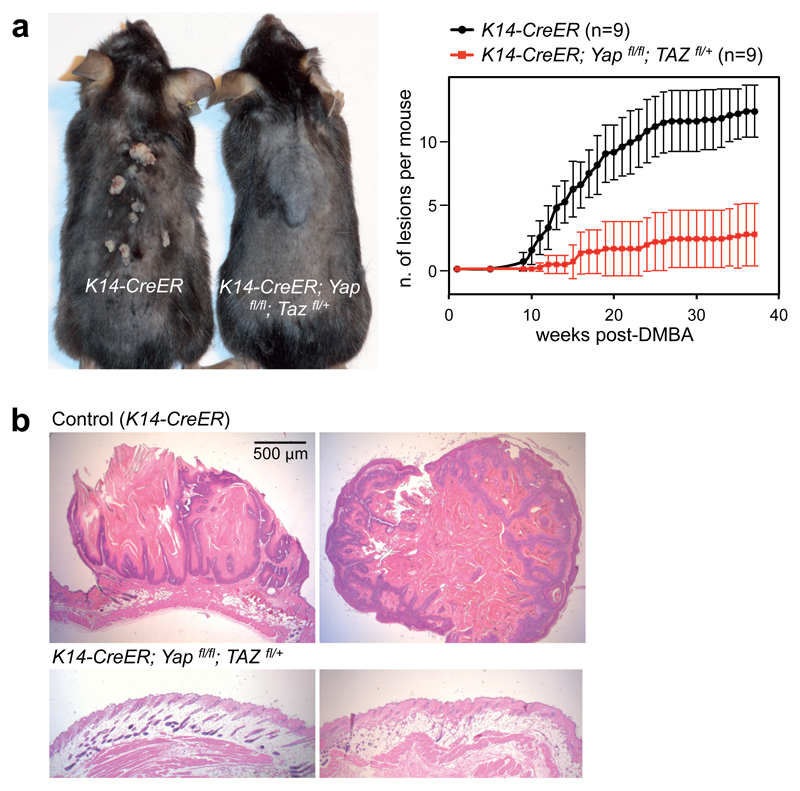
YAP/TAZ are nuclear effectors of the Hippo pathway regulating organ growth and tumorigenesis. Yet, their function as transcriptional regulators remains underinvestigated. By ChIP-seq analyses in breast cancer cells, we discovered that the YAP/TAZ transcriptional response is pervasively mediated by a dual element: TEAD factors, through which YAP/TAZ bind to DNA, co-occupying chromatin with activator protein-1 (AP-1, dimer of JUN and FOS proteins) at composite cis-regulatory elements harbouring both TEAD and AP-1 motifs. YAP/TAZ/TEAD and AP-1 form a complex that synergistically activates target genes directly involved in the control of S-phase entry and mitosis. This control occurs almost exclusively from distal enhancers that contact target promoters through chromatin looping. YAP/TAZ-induced oncogenic growth is strongly enhanced by gain of AP-1 and severely blunted by its loss. Conversely, AP-1-promoted skin tumorigenesis is prevented in YAP/TAZ conditional knockout mice. This work highlights a new layer of signalling integration, feeding on YAP/TAZ function at the chromatin level.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Similar articles
-
YAP and TAZ are transcriptional co-activators of AP-1 proteins and STAT3 during breast cellular transformation.Elife. 2021 Aug 31;10:e67312. doi: 10.7554/eLife.67312. Elife. 2021. PMID: 34463254 Free PMC article.
-
Induction of AP-1 by YAP/TAZ contributes to cell proliferation and organ growth.Genes Dev. 2020 Jan 1;34(1-2):72-86. doi: 10.1101/gad.331546.119. Epub 2019 Dec 12. Genes Dev. 2020. PMID: 31831627 Free PMC article.
-
A non-canonical repressor function of JUN restrains YAP activity and liver cancer growth.EMBO J. 2024 Oct;43(20):4578-4603. doi: 10.1038/s44318-024-00188-0. Epub 2024 Aug 29. EMBO J. 2024. PMID: 39210147 Free PMC article.
-
Multifaceted regulation and functions of YAP/TAZ in tumors (Review).Oncol Rep. 2018 Jul;40(1):16-28. doi: 10.3892/or.2018.6423. Epub 2018 May 8. Oncol Rep. 2018. PMID: 29749524 Free PMC article. Review.
-
A combat with the YAP/TAZ-TEAD oncoproteins for cancer therapy.Theranostics. 2020 Feb 18;10(8):3622-3635. doi: 10.7150/thno.40889. eCollection 2020. Theranostics. 2020. PMID: 32206112 Free PMC article. Review.
Cited by
-
YAP/TAZ enhances P-body formation to promote tumorigenesis.Elife. 2024 Jul 24;12:RP88573. doi: 10.7554/eLife.88573. Elife. 2024. PMID: 39046443 Free PMC article.
-
Plant-Based Scaffolds Modify Cellular Response to Drug and Radiation Exposure Compared to Standard Cell Culture Models.Front Bioeng Biotechnol. 2020 Aug 7;8:932. doi: 10.3389/fbioe.2020.00932. eCollection 2020. Front Bioeng Biotechnol. 2020. PMID: 32850759 Free PMC article.
-
Role of YAP Signaling in Regulation of Programmed Cell Death and Drug Resistance in Cancer.Int J Biol Sci. 2024 Jan 1;20(1):15-28. doi: 10.7150/ijbs.83586. eCollection 2024. Int J Biol Sci. 2024. PMID: 38164167 Free PMC article. Review.
-
Insights gained from computational modeling of YAP/TAZ signaling for cellular mechanotransduction.NPJ Syst Biol Appl. 2024 Aug 15;10(1):90. doi: 10.1038/s41540-024-00414-9. NPJ Syst Biol Appl. 2024. PMID: 39147782 Free PMC article. Review.
-
VAV2 signaling promotes regenerative proliferation in both cutaneous and head and neck squamous cell carcinoma.Nat Commun. 2020 Sep 22;11(1):4788. doi: 10.1038/s41467-020-18524-3. Nat Commun. 2020. PMID: 32963234 Free PMC article.
References
-
- Sudol M. Yes-associated protein (YAP65) is a proline-rich phosphoprotein that binds to the SH3 domain of the Yes proto-oncogene product. Oncogene. 1994;9:2145–2152. - PubMed
-
- Piccolo S, Dupont S, Cordenonsi M. The biology of YAP/TAZ: hippo signaling and beyond. Physiological reviews. 2014;94:1287–1312. - PubMed
-
- Harvey KF, Zhang X, Thomas DM. The Hippo pathway and human cancer. Nat Rev Cancer. 2013;13:246–257. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
