

MicroRNA-203 represses selection and expansion of oncogenic Hras transformed tumor initiating cells
- PMID: 26203562
- PMCID: PMC4536367
- DOI: 10.7554/eLife.07004
MicroRNA-203 represses selection and expansion of oncogenic Hras transformed tumor initiating cells
Abstract
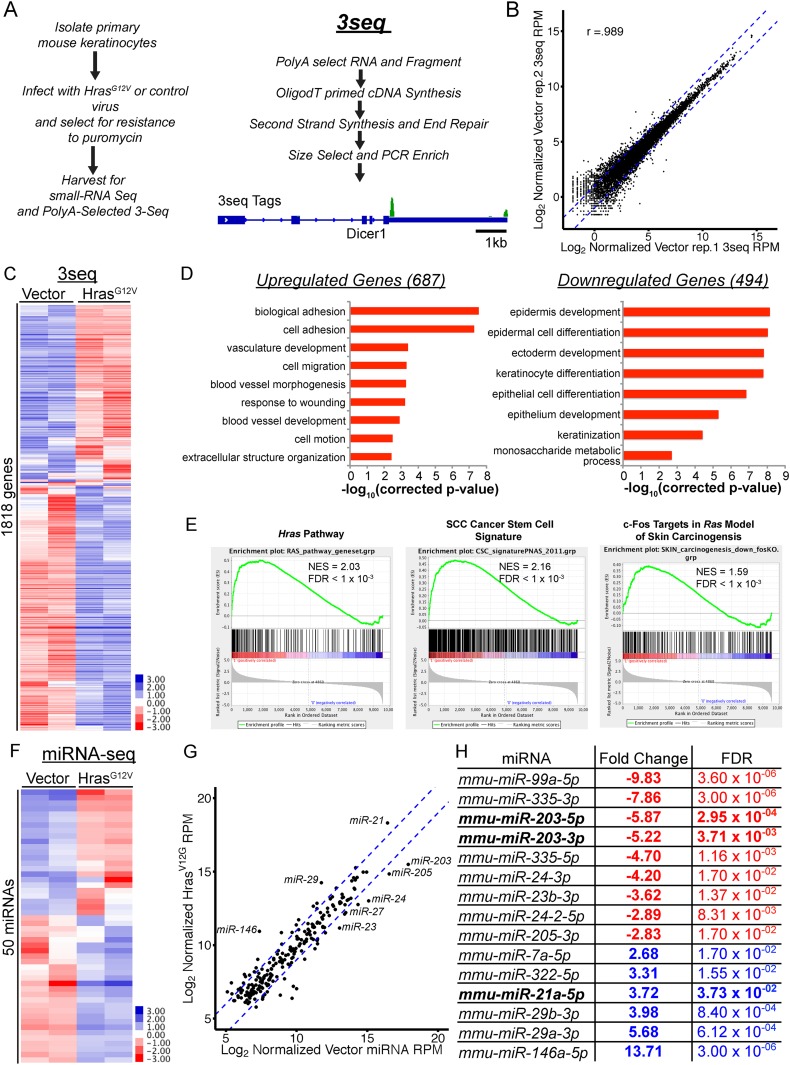
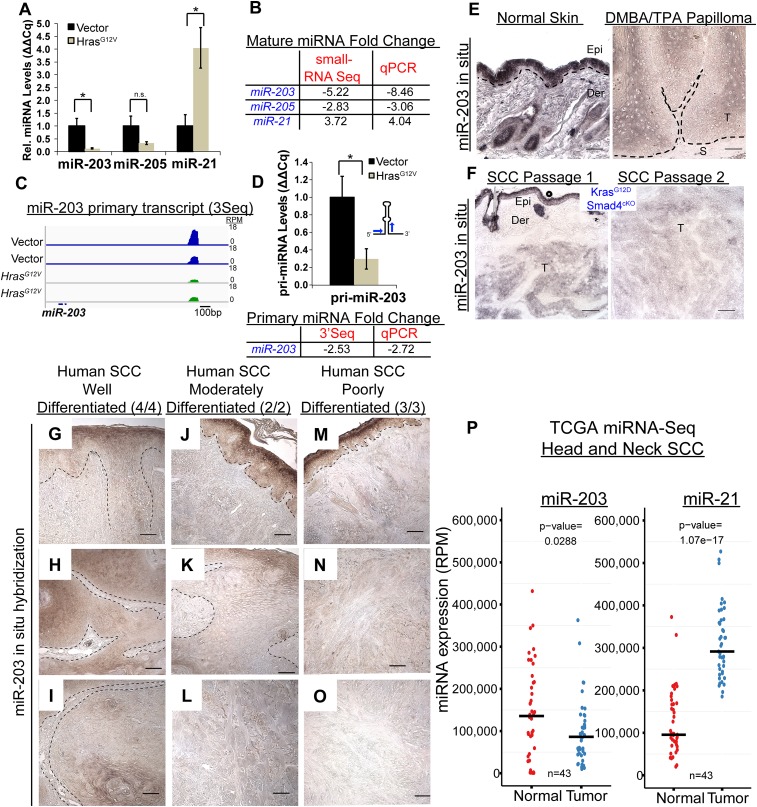
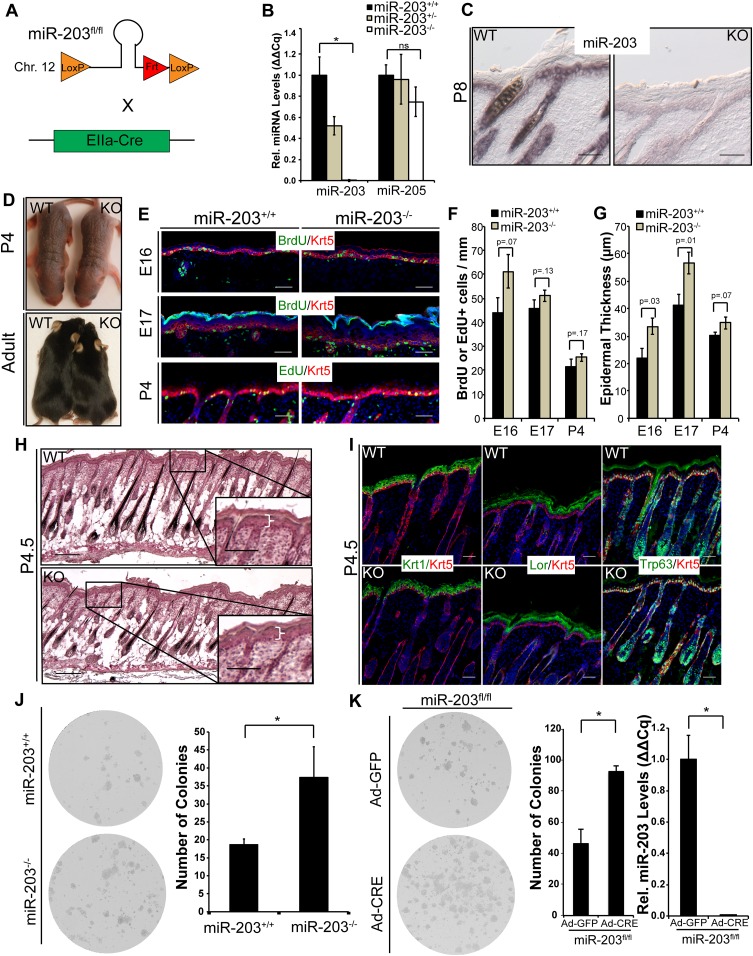
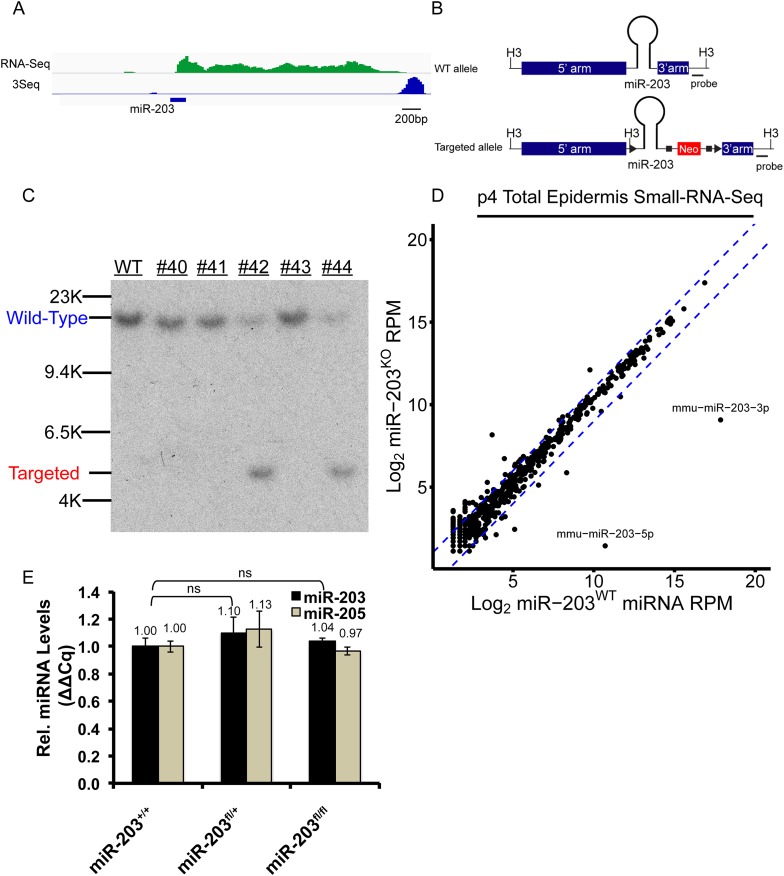
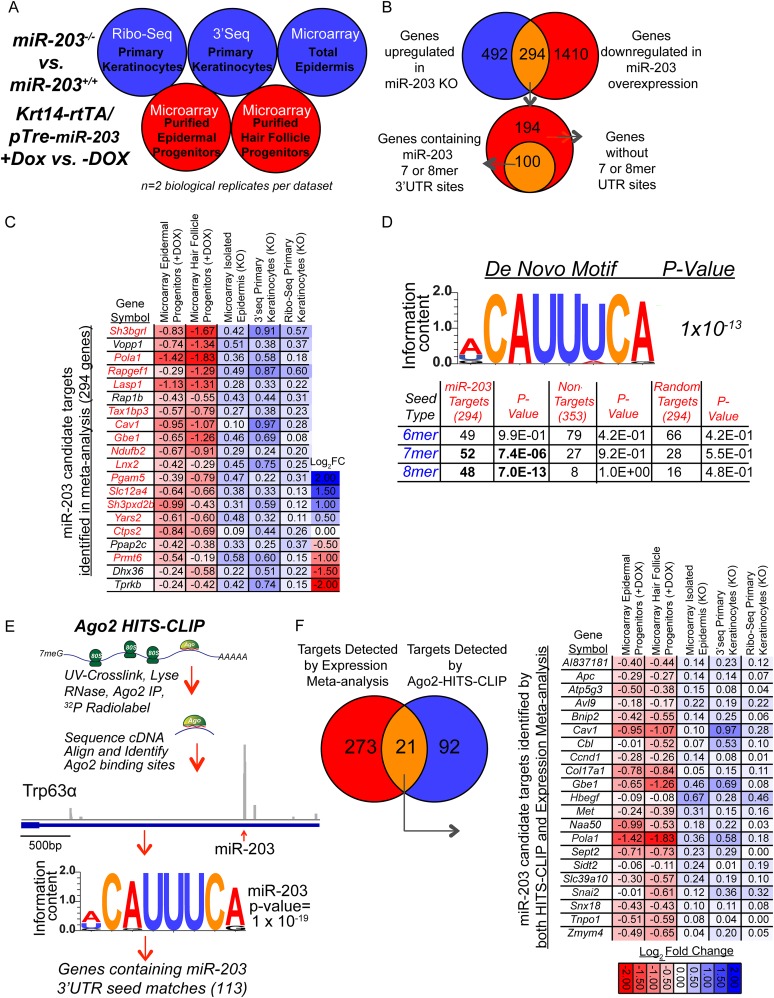
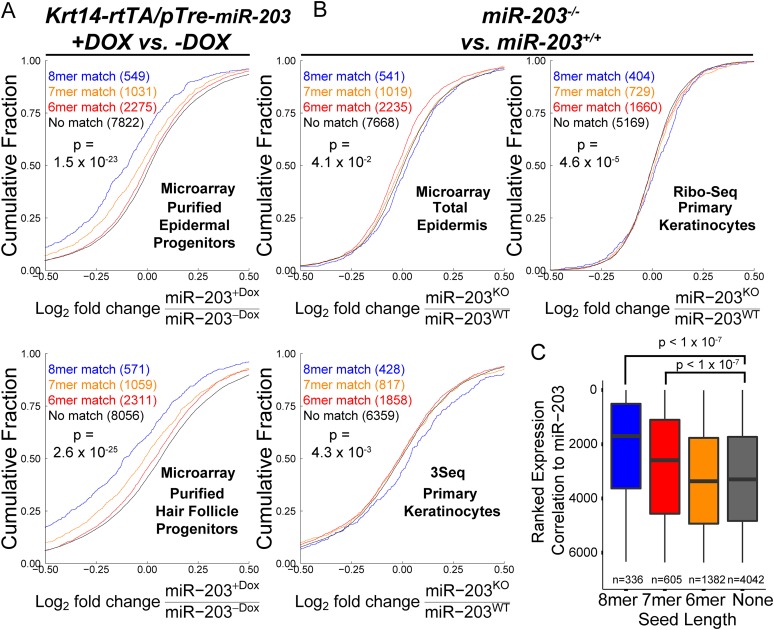
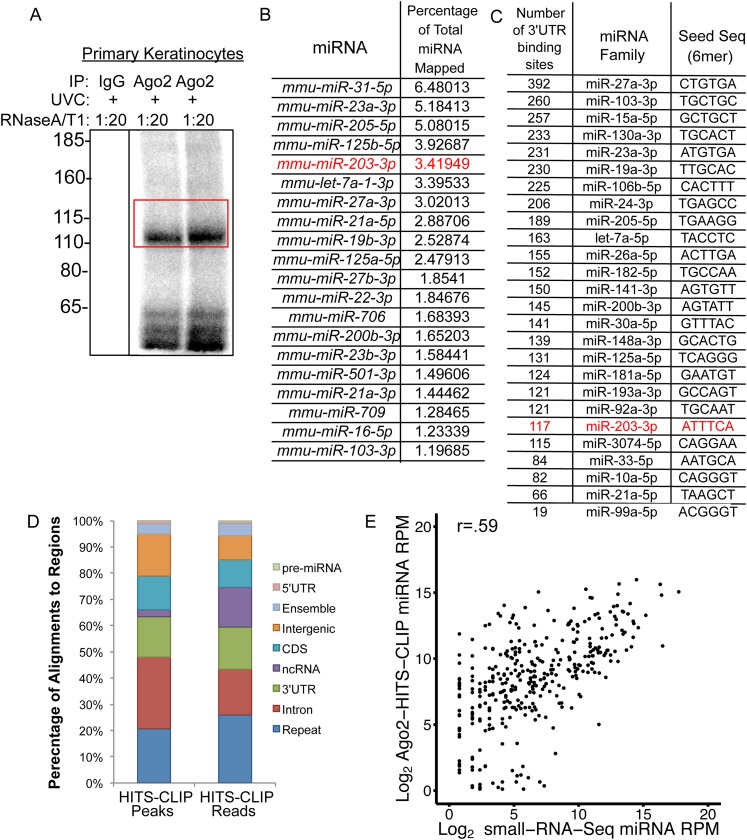
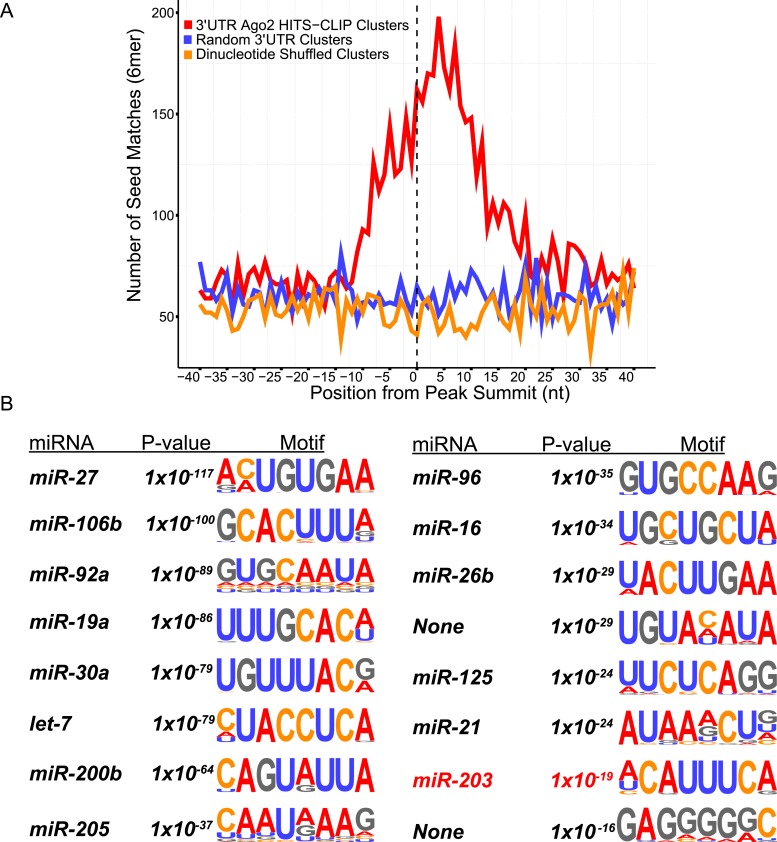
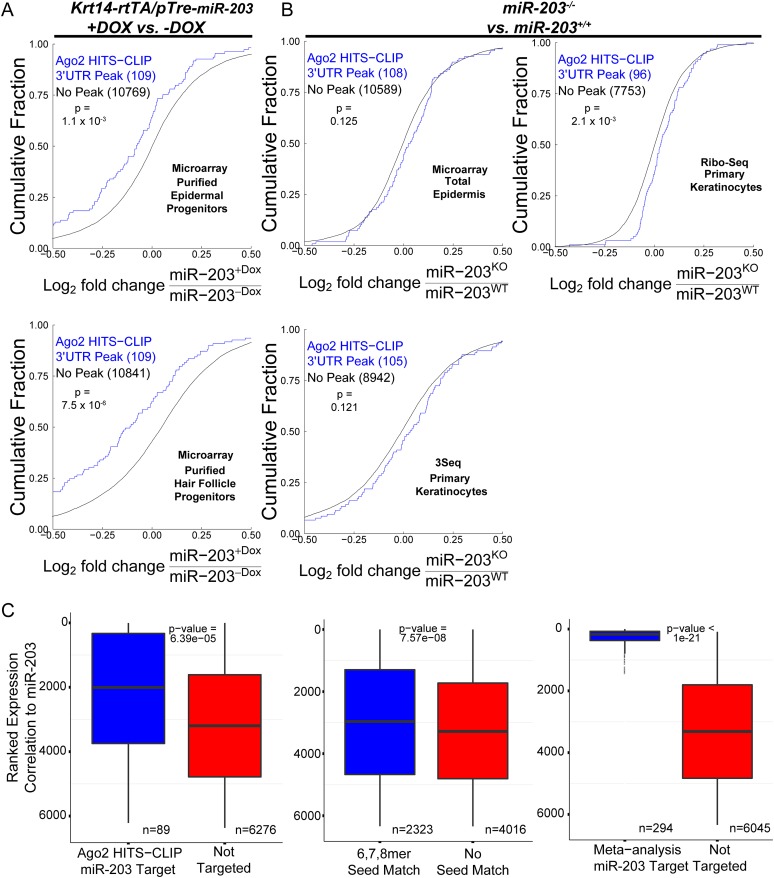
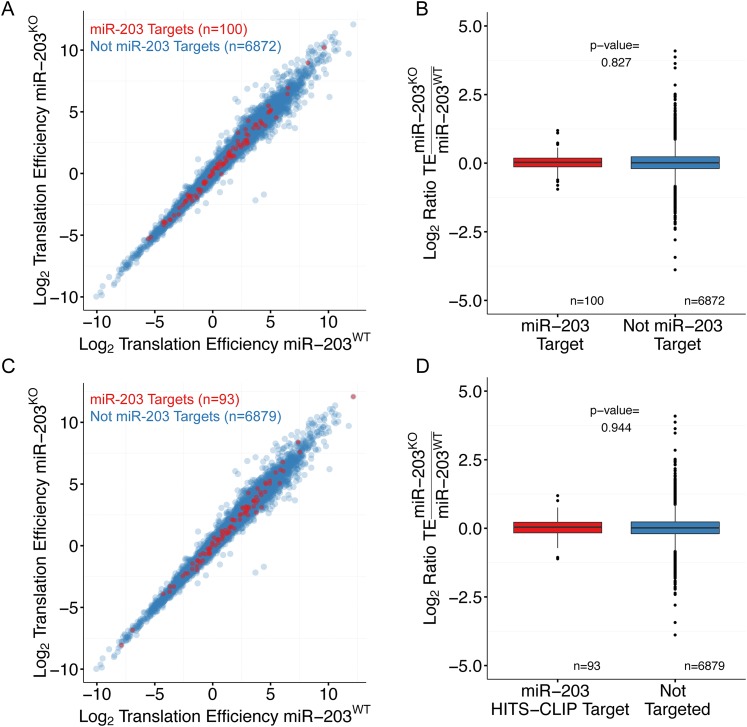
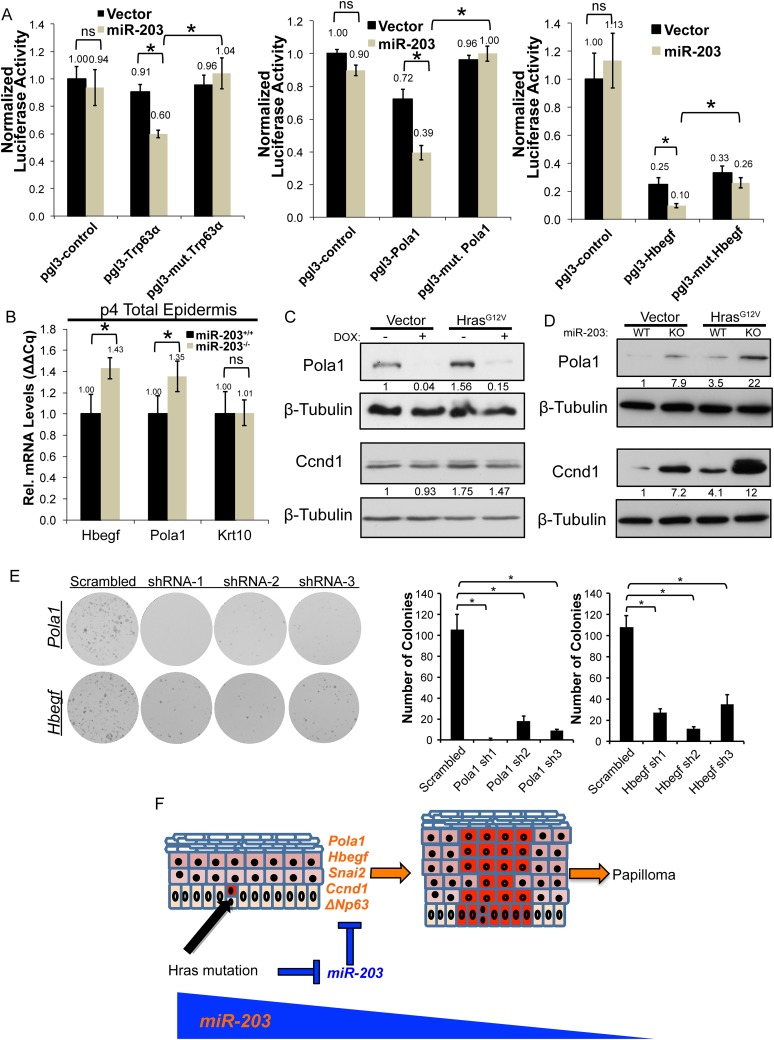
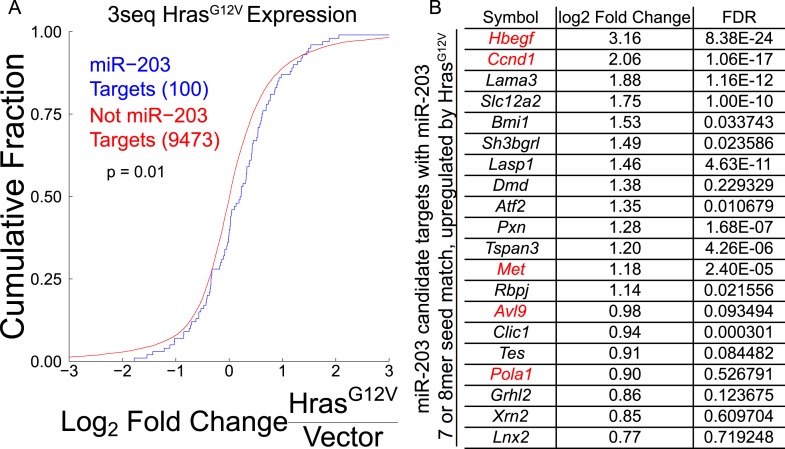
In many mouse models of skin cancer, only a few tumors typically form even though many cells competent for tumorigenesis receive the same oncogenic stimuli. These observations suggest an active selection process for tumor-initiating cells. Here, we use quantitative mRNA- and miR-Seq to determine the impact of Hras(G12V) on the transcriptome of keratinocytes. We discover that microRNA-203 is downregulated by Hras(G12V). Using a knockout mouse model, we demonstrate that loss of microRNA-203 promotes selection and expansion of tumor-initiating cells. Conversely, restoration of microRNA-203 using an inducible model potently inhibits proliferation of these cells. We comprehensively identify microRNA-203 targets required for Hras-initiated tumorigenesis. These targets include critical regulators of the Ras pathway and essential genes required for cell division. This study establishes a role for the loss of microRNA-203 in promoting selection and expansion of Hras mutated cells and identifies a mechanism through which microRNA-203 antagonizes Hras-mediated tumorigenesis.
Keywords: Hras; developmental biology; human biology; medicine; microRNA; mouse; skin cancer; stem cells; tumor suppressor.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
Similar articles
-
Reduced HRAS G12V-Driven Tumorigenesis of Cell Lines Expressing KRAS C118S.PLoS One. 2015 Apr 22;10(4):e0123918. doi: 10.1371/journal.pone.0123918. eCollection 2015. PLoS One. 2015. PMID: 25902334 Free PMC article.
-
A critical role of MYC for transformation of human cells by HPV16 E6E7 and oncogenic HRAS.Carcinogenesis. 2012 Apr;33(4):910-7. doi: 10.1093/carcin/bgs104. Epub 2012 Feb 15. Carcinogenesis. 2012. PMID: 22345164
-
The role of oncogenic Ras in human skin tumorigenesis depends on the clonogenic potential of the founding keratinocytes.J Cell Sci. 2016 Mar 1;129(5):1003-17. doi: 10.1242/jcs.176842. Epub 2016 Jan 21. J Cell Sci. 2016. PMID: 26795563
-
Transforming growth factor-beta and microRNA:mRNA regulatory networks in epithelial plasticity.Cells Tissues Organs. 2007;185(1-3):157-61. doi: 10.1159/000101316. Cells Tissues Organs. 2007. PMID: 17587821 Review.
-
Molecular and cellular basis for tumor promotion in mouse skin.Princess Takamatsu Symp. 1983;14:315-26. Princess Takamatsu Symp. 1983. PMID: 6394593 Review.
Cited by
-
miR-22 promotes stem cell traits via activating Wnt/β-catenin signaling in cutaneous squamous cell carcinoma.Oncogene. 2021 Sep;40(39):5799-5813. doi: 10.1038/s41388-021-01973-5. Epub 2021 Aug 3. Oncogene. 2021. PMID: 34345013 Free PMC article.
-
The microRNA-200 family coordinately regulates cell adhesion and proliferation in hair morphogenesis.J Cell Biol. 2018 Jun 4;217(6):2185-2204. doi: 10.1083/jcb.201708173. Epub 2018 Mar 30. J Cell Biol. 2018. PMID: 29602800 Free PMC article.
-
Metazoan MicroRNAs.Cell. 2018 Mar 22;173(1):20-51. doi: 10.1016/j.cell.2018.03.006. Cell. 2018. PMID: 29570994 Free PMC article. Review.
-
Cancer Stem Cells in Squamous Cell Carcinoma.J Invest Dermatol. 2017 Jan;137(1):31-37. doi: 10.1016/j.jid.2016.07.033. Epub 2016 Nov 24. J Invest Dermatol. 2017. PMID: 27638386 Free PMC article. Review.
-
Functional robustness of adult spermatogonial stem cells after induction of hyperactive Hras.PLoS Genet. 2019 May 3;15(5):e1008139. doi: 10.1371/journal.pgen.1008139. eCollection 2019 May. PLoS Genet. 2019. PMID: 31050682 Free PMC article.
References
-
- Agrawal N, Frederick MJ, Pickering CR, Bettegowda C, Chang K, Li RJ, Fakhry C, Xie TX, Zhang J, Wang J, Zhang N, El-Naggar AK, Jasser SA, Weinstein JN, Treviño L, Drummond JA, Muzny DM, Wu Y, Wood LD, Hruban RH, Westra WH, Koch WM, Califano JA, Gibbs RA, Sidransky D, Vogelstein B, Velculescu VE, Papadopoulos N, Wheeler DA, Kinzler KW, Myers JN. Exome sequencing of head and neck squamous cell carcinoma reveals inactivating mutations in NOTCH1. Science. 2011;333:1154–1157. doi: 10.1126/science.1206923. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
