

Differential recruitment of UBQLN2 to nuclear inclusions in the polyglutamine diseases HD and SCA3
- PMID: 26141599
- PMCID: PMC4642276
- DOI: 10.1016/j.nbd.2015.06.017
Differential recruitment of UBQLN2 to nuclear inclusions in the polyglutamine diseases HD and SCA3
Abstract
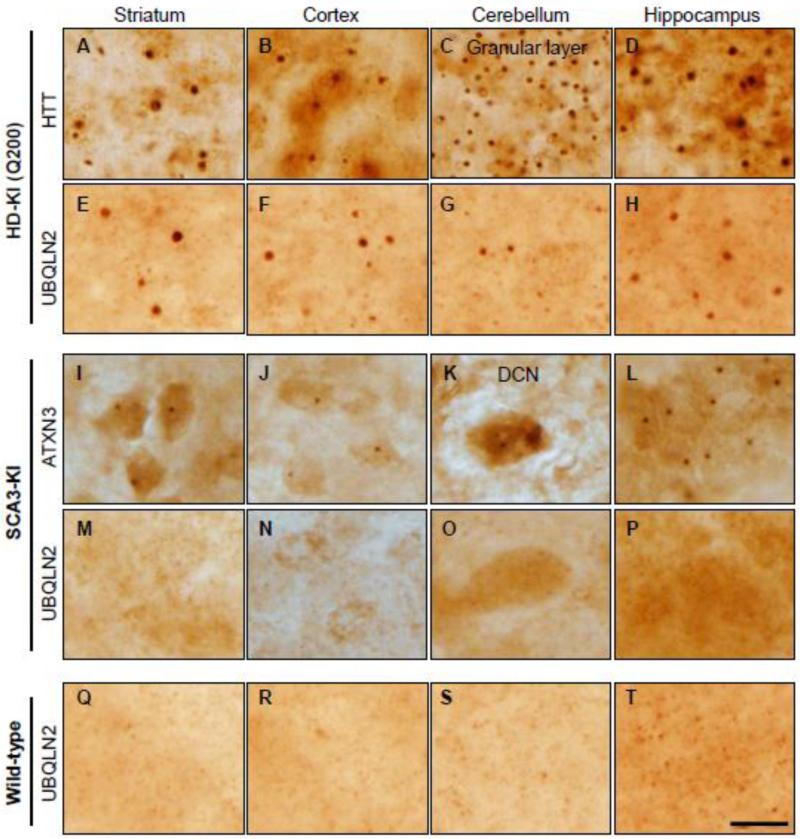
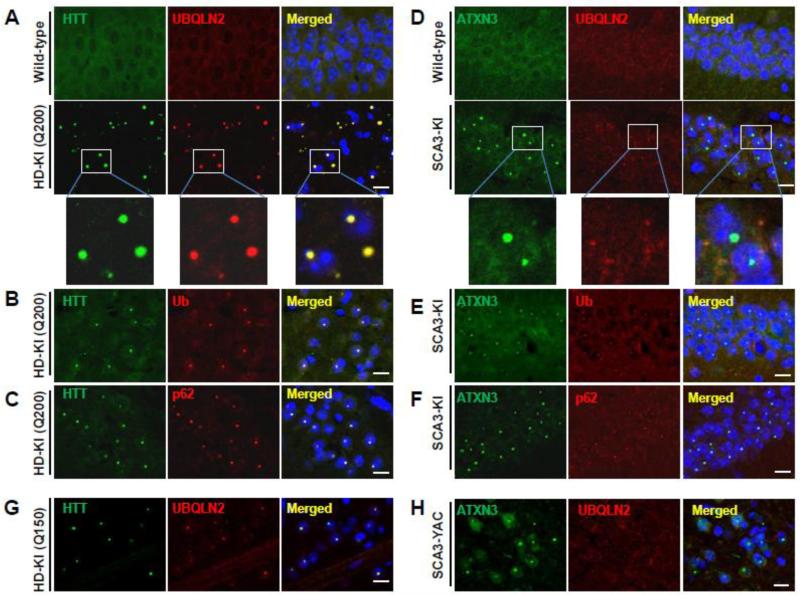
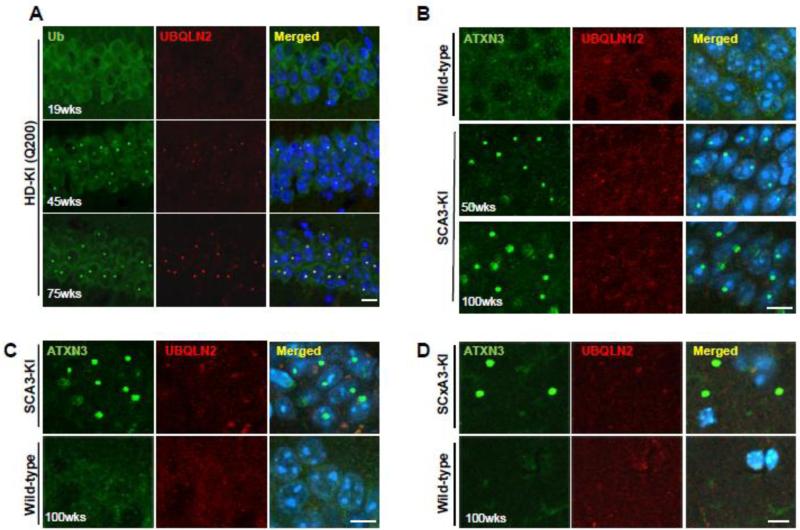
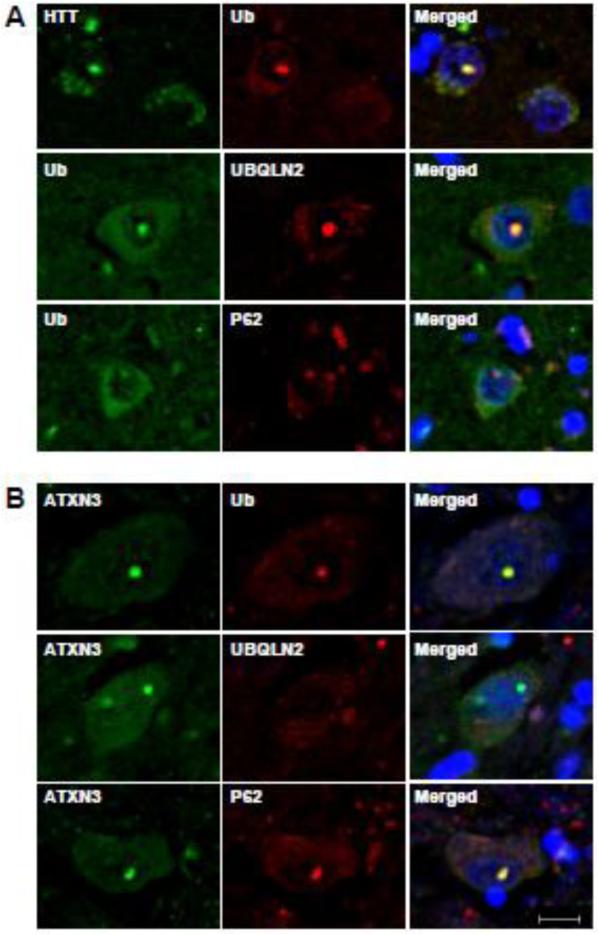
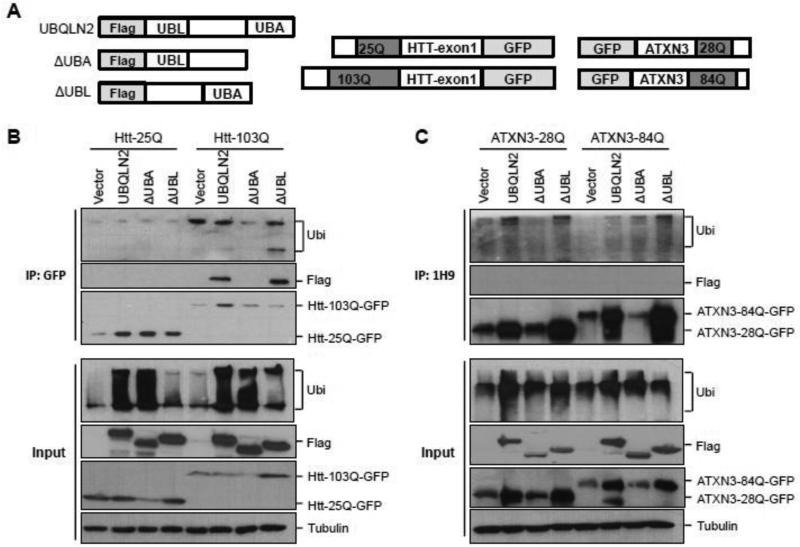
Accumulation of mutant polyglutamine proteins in intraneuronal inclusions is a hallmark of polyglutamine diseases. Impairment of protein clearance systems and sequestration of clearance-related proteins into inclusions occur in many protein folding diseases, including polyglutamine diseases. The ubiquitin-binding and proteasome adaptor protein UBQLN2 participates in protein homeostasis and localizes to inclusions in various neurodegenerative diseases. Employing mouse models and human brain tissue of Huntington's disease (HD) and spinocerebellar ataxia type 3 (SCA3), we show that UBQLN2 is selectively recruited to inclusions in HD but not SCA3. Consistent with this result, in a cell-based system mutant HTT interacts with UBQLN2 through the UBA domain while the SCA3 disease protein ATXN3, a deubiquitinating enzyme, does not interact with UBQLN2. Differential recruitment of UBQLN2 to aggregates in HD and SCA3 underscores the heterogeneity of inclusions in polyglutamine diseases and suggests that components of neuronal protein quality control may be differentially perturbed in distinct polyQ diseases.
Keywords: Huntington's disease; Polyglutamine; SCA3; UBA; UBL; UBQLN2.
Copyright © 2015. Published by Elsevier Inc.
Figures
Similar articles
-
Ubiquilin-2 differentially regulates polyglutamine disease proteins.Hum Mol Genet. 2020 Aug 29;29(15):2596-2610. doi: 10.1093/hmg/ddaa152. Hum Mol Genet. 2020. PMID: 32681165 Free PMC article.
-
PolyQ-expanded huntingtin and ataxin-3 sequester ubiquitin adaptors hHR23B and UBQLN2 into aggregates via conjugated ubiquitin.FASEB J. 2018 Jun;32(6):2923-2933. doi: 10.1096/fj.201700801RR. Epub 2018 Jan 11. FASEB J. 2018. PMID: 29401586
-
A new humanized ataxin-3 knock-in mouse model combines the genetic features, pathogenesis of neurons and glia and late disease onset of SCA3/MJD.Neurobiol Dis. 2015 Jan;73:174-88. doi: 10.1016/j.nbd.2014.09.020. Epub 2014 Oct 7. Neurobiol Dis. 2015. PMID: 25301414
-
Toward therapeutic targets for SCA3: Insight into the role of Machado-Joseph disease protein ataxin-3 in misfolded proteins clearance.Prog Neurobiol. 2015 Sep;132:34-58. doi: 10.1016/j.pneurobio.2015.06.004. Epub 2015 Jun 27. Prog Neurobiol. 2015. PMID: 26123252 Review.
-
Autophagy in Spinocerebellar Ataxia Type 3: From Pathogenesis to Therapeutics.Int J Mol Sci. 2023 Apr 17;24(8):7405. doi: 10.3390/ijms24087405. Int J Mol Sci. 2023. PMID: 37108570 Free PMC article. Review.
Cited by
-
Profilin reduces aggregation and phase separation of huntingtin N-terminal fragments by preferentially binding to soluble monomers and oligomers.J Biol Chem. 2018 Mar 9;293(10):3734-3746. doi: 10.1074/jbc.RA117.000357. Epub 2018 Jan 22. J Biol Chem. 2018. PMID: 29358329 Free PMC article.
-
UBQLN Family Members Regulate MYC in Lung Adenocarcinoma Cells.Cancers (Basel). 2023 Jun 28;15(13):3389. doi: 10.3390/cancers15133389. Cancers (Basel). 2023. PMID: 37444499 Free PMC article.
-
Peripheral sequestration of huntingtin delays neuronal death and depends on N-terminal ubiquitination.Commun Biol. 2024 Aug 18;7(1):1014. doi: 10.1038/s42003-024-06733-1. Commun Biol. 2024. PMID: 39155290 Free PMC article.
-
Carboxyl Terminus of Hsp70-Interacting Protein Is Increased in Serum and Cerebrospinal Fluid of Patients With Spinocerebellar Ataxia Type 3.Front Neurol. 2019 Oct 15;10:1094. doi: 10.3389/fneur.2019.01094. eCollection 2019. Front Neurol. 2019. PMID: 31749756 Free PMC article.
-
Coaggregation of polyglutamine (polyQ) proteins is mediated by polyQ-tract interactions and impairs cellular proteostasis.Acta Biochim Biophys Sin (Shanghai). 2023 May 11;55(5):736-748. doi: 10.3724/abbs.2023081. Acta Biochim Biophys Sin (Shanghai). 2023. PMID: 37171184 Free PMC article.
References
-
- Cemal CK, et al. YAC transgenic mice carrying pathological alleles of the MJD1 locus exhibit a mild and slowly progressive cerebellar deficit. Hum Mol Genet. 2002;11:1075–94. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
