

Mature T cell responses are controlled by microRNA-142
- PMID: 26098216
- PMCID: PMC4563679
- DOI: 10.1172/JCI78753
Mature T cell responses are controlled by microRNA-142
Abstract
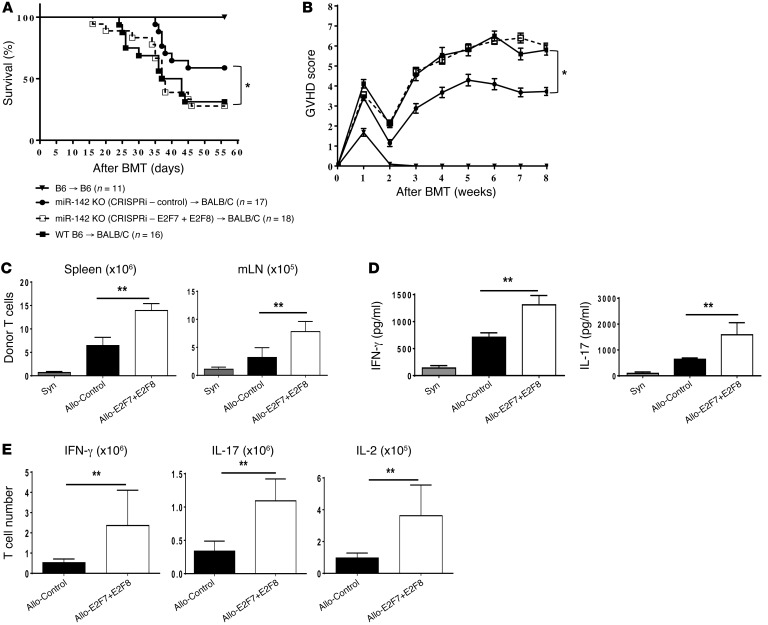
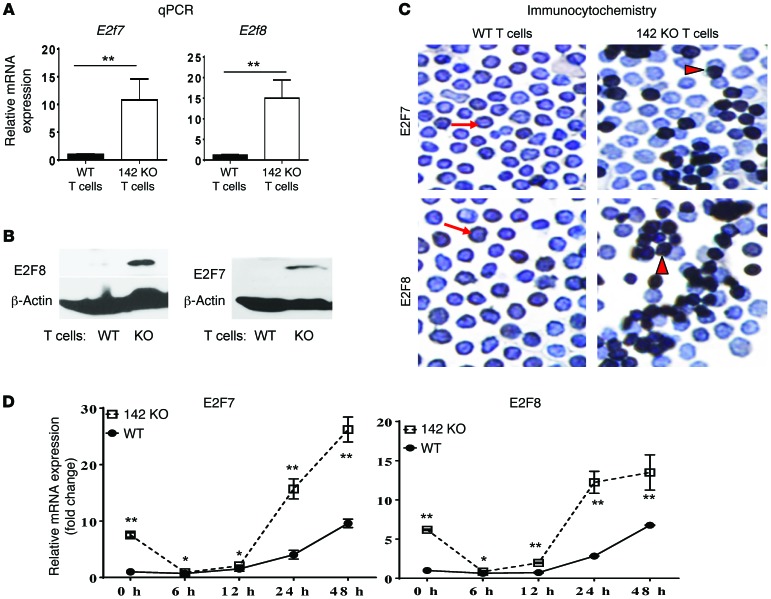
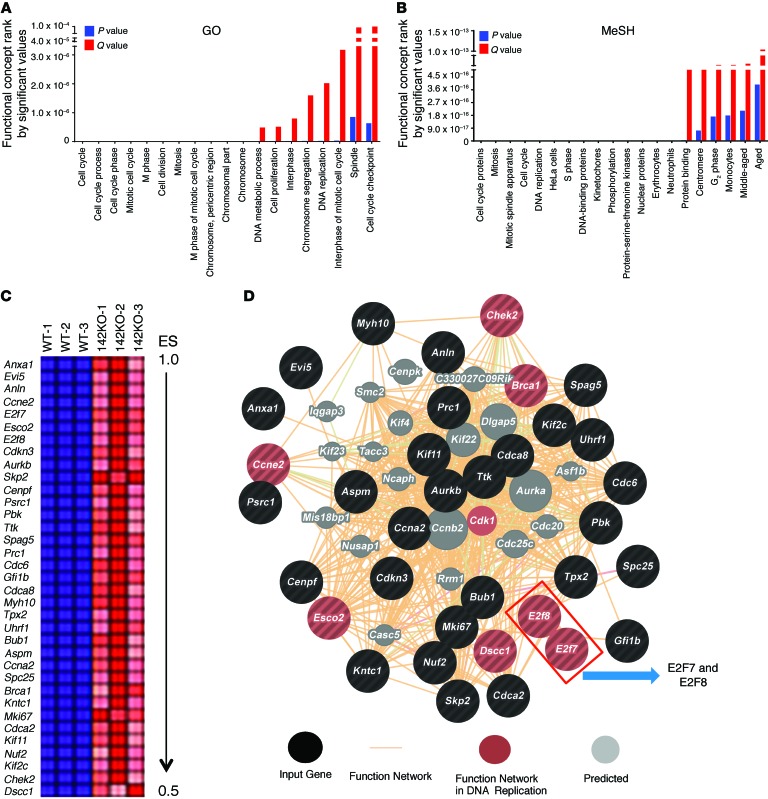
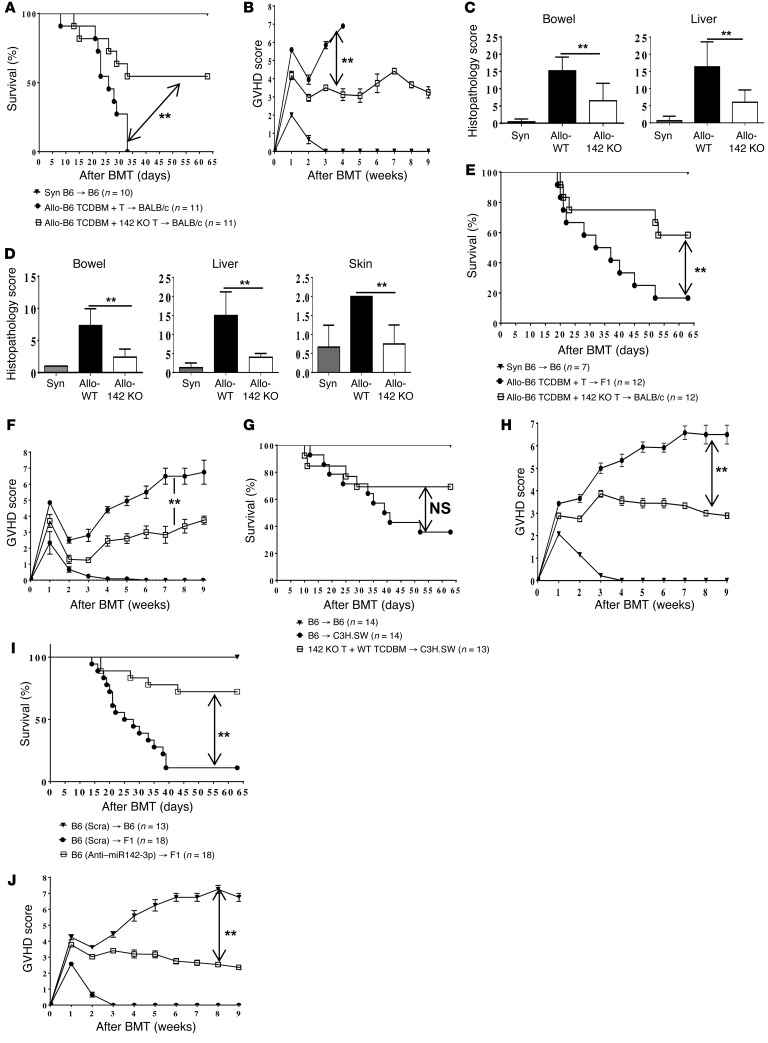
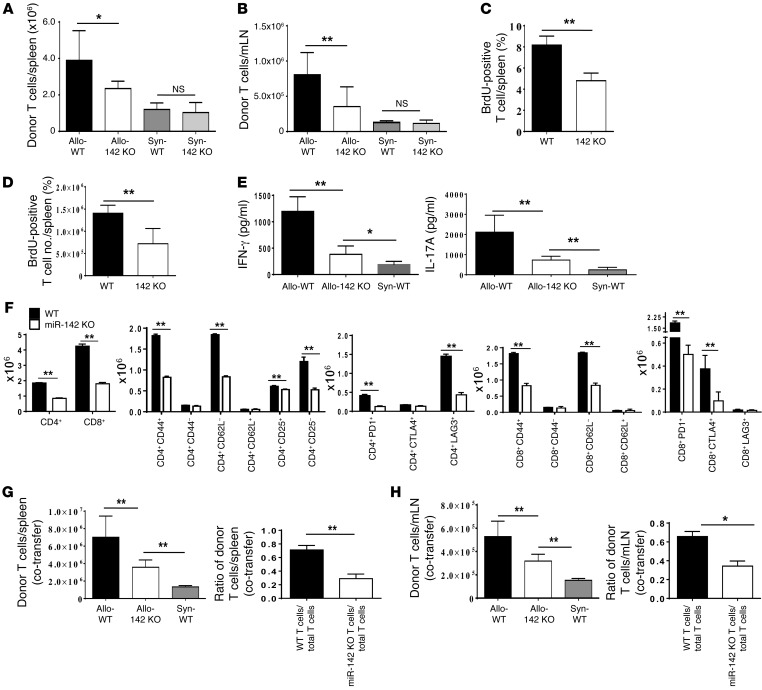
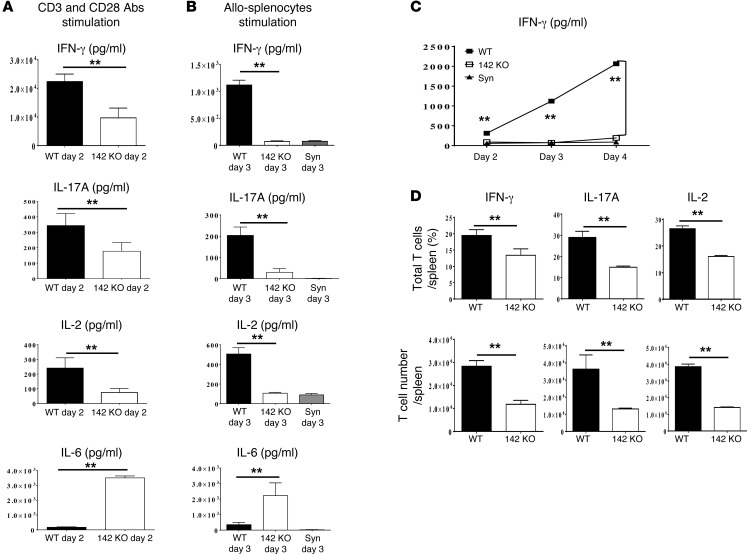
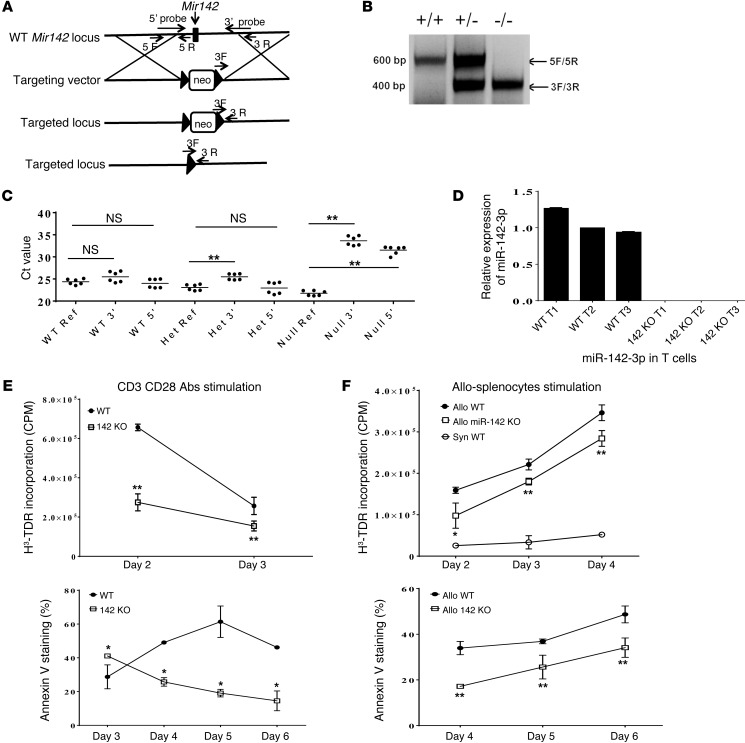
T cell proliferation is critical for immune responses; however, the molecular mechanisms that mediate the proliferative response are poorly understood. MicroRNAs (miRs) regulate various molecular processes, including development and function of the immune system. Here, utilizing multiple complementary genetic and molecular approaches, we investigated the contribution of a hematopoietic-specific miR, miR-142, in regulating T cell responses. T cell development was not affected in animals with a targeted deletion of Mir142; however, T cell proliferation was markedly reduced following stimulation both in vitro and in multiple murine models of graft-versus-host disease (GVHD). miR-142-deficient T cells demonstrated substantial cell-cycling defects, and microarray and bioinformatics analyses revealed upregulation of genes involved in cell cycling. Moreover, 2 predicted miR-142 target genes, the atypical E2F transcription factors E2f7 and E2f8, were most highly upregulated in miR-142-deficient cells. Clustered regularly interspaced short palindromic repeat interference-mediated (CRISPRi-mediated) silencing of E2F7 and E2F8 in miR-142-deficient T cells ameliorated cell-cycling defects and reduced GVHD, and overexpression of these factors in WT T cells inhibited the proliferative response. Together, these results identify a link between hematopoietic-specific miR-142 and atypical E2F transcription factors in the regulation of mature T cell cycling and suggest that targeting this interaction may be relevant for mitigating GVHD.
Figures

Similar articles
-
MicroRNA-17-92 controls T-cell responses in graft-versus-host disease and leukemia relapse in mice.Blood. 2015 Sep 10;126(11):1314-23. doi: 10.1182/blood-2015-02-627356. Epub 2015 Jul 2. Blood. 2015. PMID: 26138686 Free PMC article.
-
Synergistic functions of E2F7 and E2F8 are critical to suppress stress-induced skin cancer.Oncogene. 2017 Feb 9;36(6):829-839. doi: 10.1038/onc.2016.251. Epub 2016 Jul 25. Oncogene. 2017. PMID: 27452520 Free PMC article.
-
Interleukin 21 blockade modulates activated T- and B-cell homeostasis via B-cell activating factor pathway-mediated inhibition in a murine model of acute graft-versus-host disease.Exp Hematol. 2015 Jan;43(1):23-31.e1-2. doi: 10.1016/j.exphem.2014.09.005. Epub 2014 Sep 20. Exp Hematol. 2015. PMID: 25246268
-
MicroRNA in T-Cell Development and T-Cell Mediated Acute Graft-Versus-Host Disease.Front Immunol. 2018 May 7;9:992. doi: 10.3389/fimmu.2018.00992. eCollection 2018. Front Immunol. 2018. PMID: 29867969 Free PMC article. Review.
-
The Role of MicroRNAs in Myeloid Cells during Graft-versus-Host Disease.Front Immunol. 2018 Jan 23;9:4. doi: 10.3389/fimmu.2018.00004. eCollection 2018. Front Immunol. 2018. PMID: 29410665 Free PMC article. Review.
Cited by
-
Patterns of immune infiltration in lung adenocarcinoma revealed a prognosis-associated microRNA-mast cells network.Hum Cell. 2020 Jan;33(1):205-219. doi: 10.1007/s13577-019-00300-1. Epub 2019 Dec 20. Hum Cell. 2020. PMID: 31863291
-
Insights into mechanisms of graft-versus-host disease through humanised mouse models.Biosci Rep. 2022 Sep 30;42(9):BSR20211986. doi: 10.1042/BSR20211986. Biosci Rep. 2022. PMID: 35993192 Free PMC article. Review.
-
MicroRNA-142 Is Critical for the Homeostasis and Function of Type 1 Innate Lymphoid Cells.Immunity. 2019 Sep 17;51(3):479-490.e6. doi: 10.1016/j.immuni.2019.06.016. Epub 2019 Aug 8. Immunity. 2019. PMID: 31402259 Free PMC article.
-
A gain-of-function mutation in microRNA 142 is sufficient to cause the development of T-cell leukemia in mice.Cancer Sci. 2023 Jul;114(7):2821-2834. doi: 10.1111/cas.15794. Epub 2023 Apr 3. Cancer Sci. 2023. PMID: 36945113 Free PMC article.
-
Integrative genomic analysis of peritoneal malignant mesothelioma: understanding a case with extraordinary chemotherapy response.Cold Spring Harb Mol Case Stud. 2019 Apr 1;5(2):a003566. doi: 10.1101/mcs.a003566. Print 2019 Apr. Cold Spring Harb Mol Case Stud. 2019. PMID: 30862609 Free PMC article.
References
-
- Haasch D, et al. T cell activation induces a noncoding RNA transcript sensitive to inhibition by immunosuppressant drugs and encoded by the proto-oncogene, BIC. Cell Immunol. 2002;217(1–2):78–86. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
