

Gibberellin Promotes Shoot Branching in the Perennial Woody Plant Jatropha curcas
- PMID: 26076970
- PMCID: PMC4523387
- DOI: 10.1093/pcp/pcv089
Gibberellin Promotes Shoot Branching in the Perennial Woody Plant Jatropha curcas
Abstract
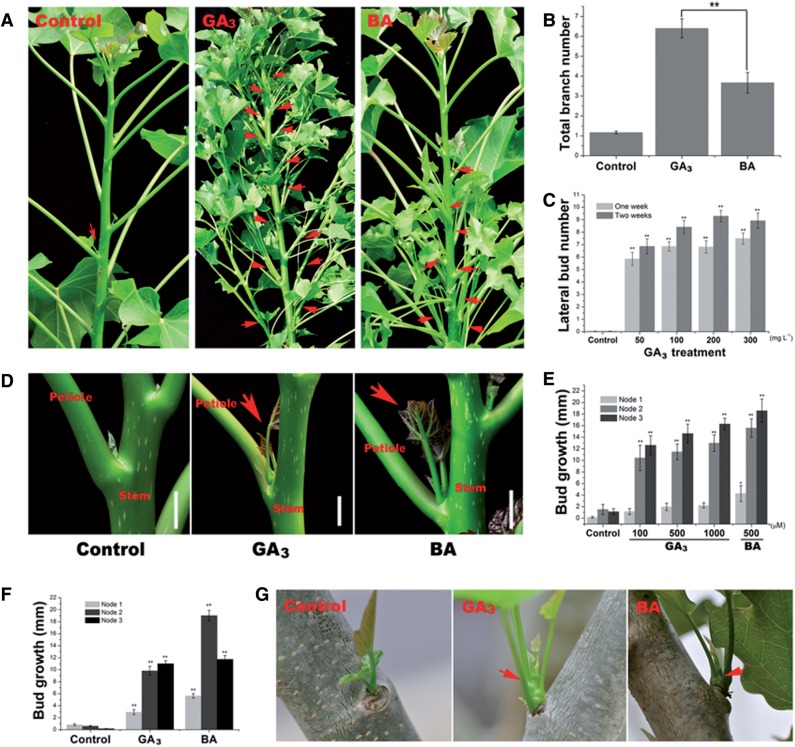
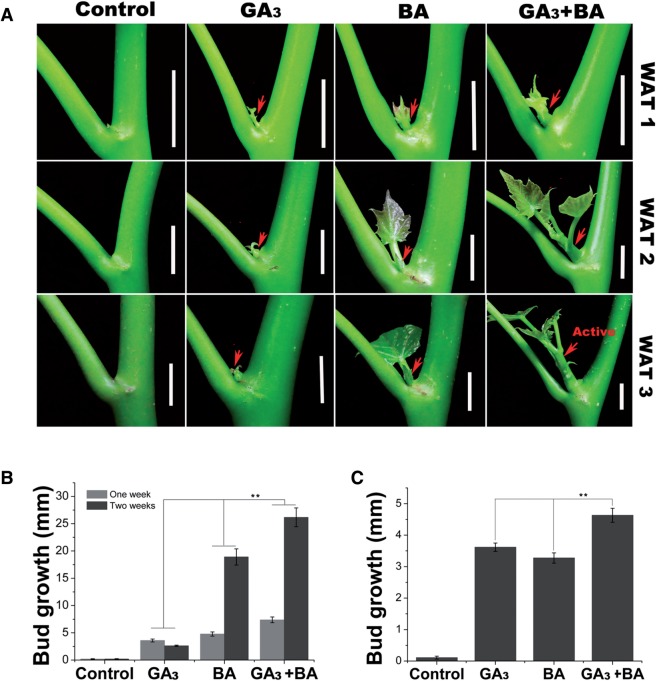
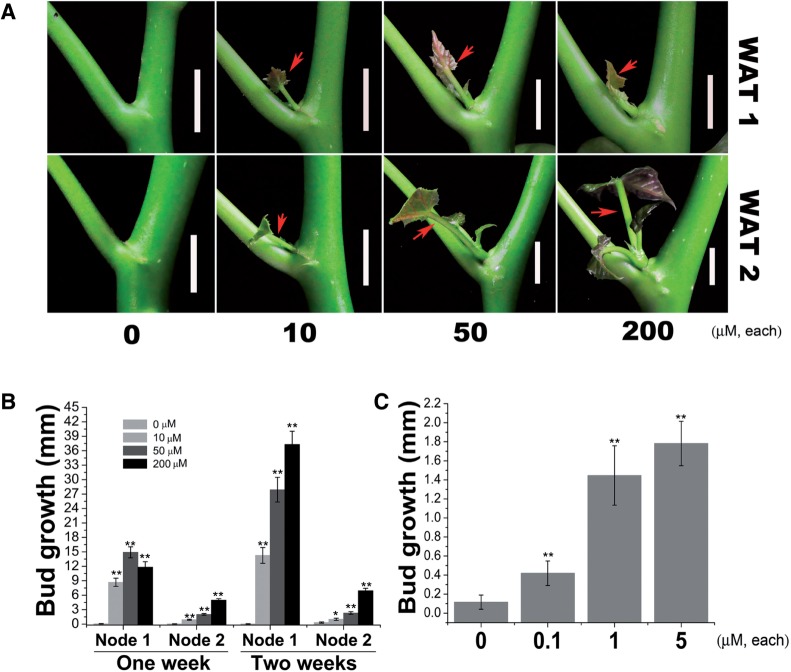
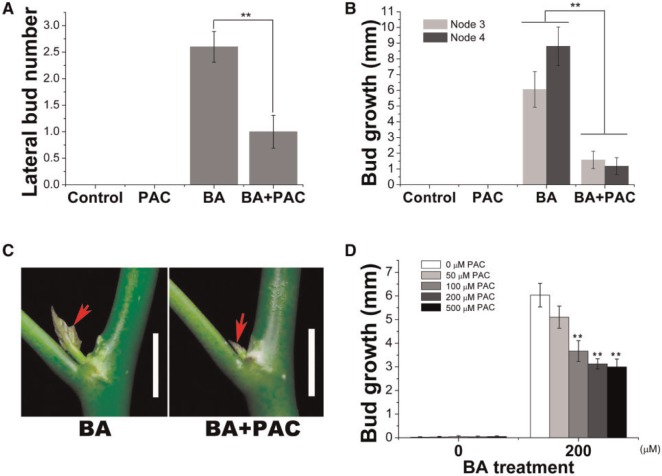
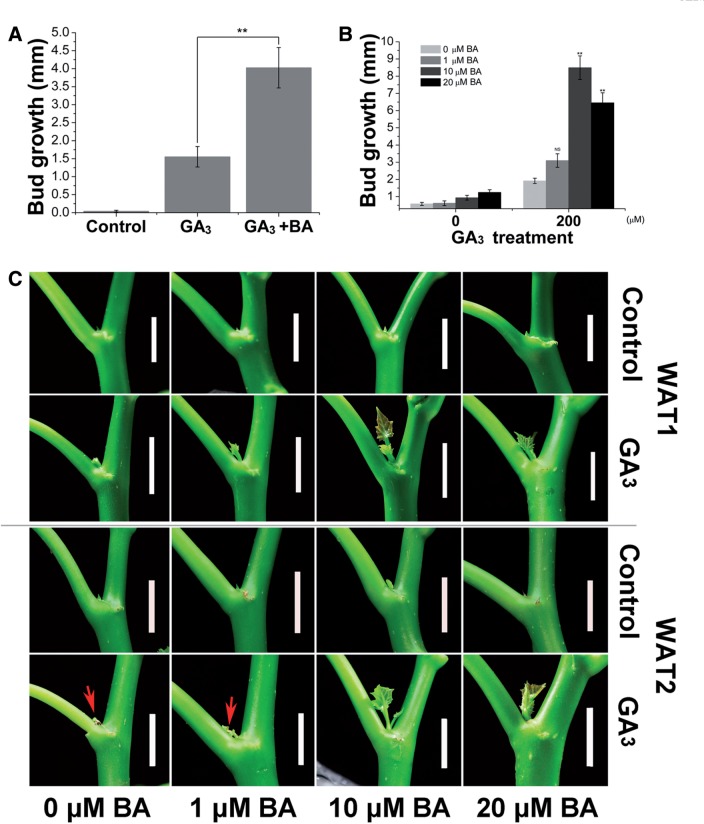
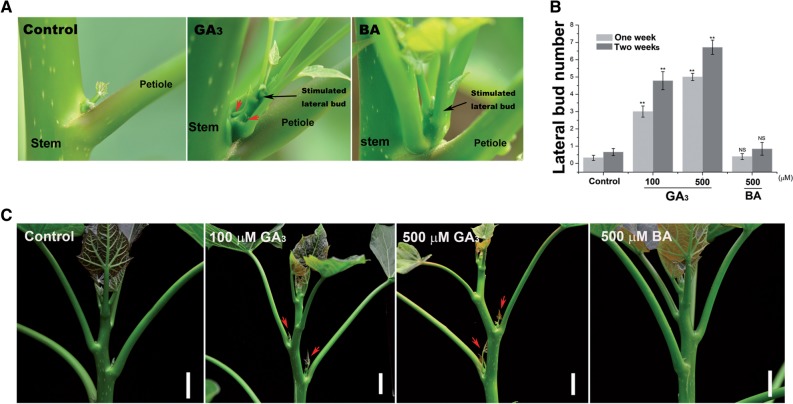
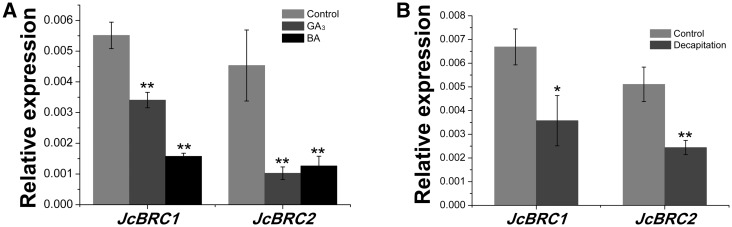
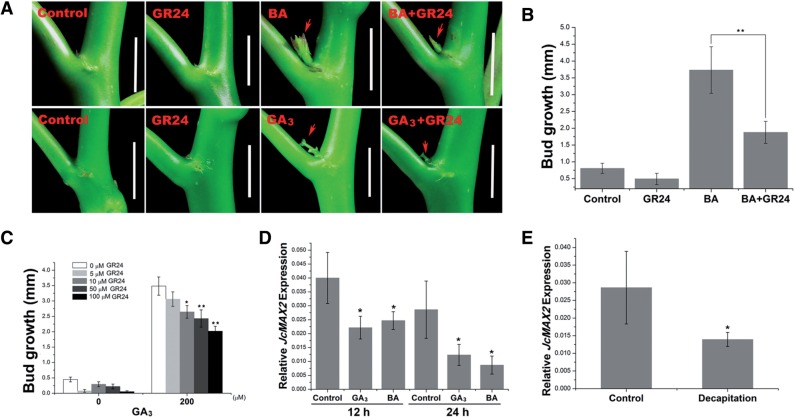
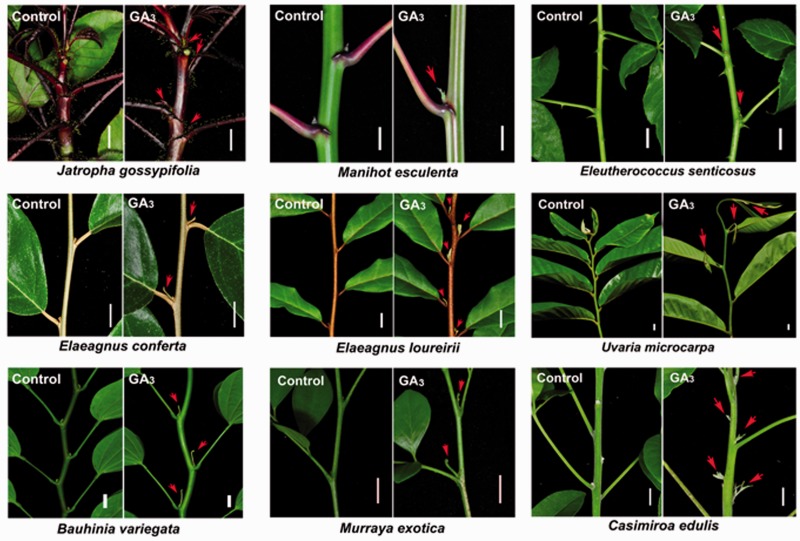
Strigolactone (SL), auxin and cytokinin (CK) interact to regulate shoot branching. CK has long been considered to be the only key phytohormone to promote lateral bud outgrowth. Here we report that gibberellin also acts as a positive regulator in the control of shoot branching in the woody plant Jatropha curcas. We show that gibberellin and CK synergistically promote lateral bud outgrowth, and that both hormones influence the expression of putative branching regulators, J. curcas BRANCHED1 and BRANCHED2, which are key transcription factors maintaining bud dormancy. Moreover, treatment with paclobutrazol, an inhibitor of de novo gibberellin biosynthesis, significantly reduced the promotion of bud outgrowth by CK, suggesting that gibberellin is required for CK-mediated axillary bud outgrowth. In addition, SL, a plant hormone involved in the repression of shoot branching, acted antagonistically to both gibberellin and CK in the control of lateral bud outgrowth. Consistent with this, the expression of JcMAX2, a J. curcas homolog of Arabidopsis MORE AXILLARY GROWTH 2 encoding an F-box protein in the SL signaling pathway, was repressed by gibberellin and CK treatment. We also provide physiological evidence that gibberellin also induces shoot branching in many other trees, such as papaya, indicating that a more complicated regulatory network occurs in the control of shoot branching in some perennial woody plants.
Keywords: Axillary bud; Bud outgrowth; Cytokinin; Gibberellin; Shoot branching; Strigolactone.
© The Author 2015. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.
Figures

Similar articles
-
Comparative transcriptome analysis of axillary buds in response to the shoot branching regulators gibberellin A3 and 6-benzyladenine in Jatropha curcas.Sci Rep. 2017 Sep 12;7(1):11417. doi: 10.1038/s41598-017-11588-0. Sci Rep. 2017. PMID: 28900192 Free PMC article.
-
Genome-Wide Identification and Expression Analysis of Cytokinin Response Regulator (RR) Genes in the Woody Plant Jatropha curcas and Functional Analysis of JcRR12 in Arabidopsis.Int J Mol Sci. 2022 Sep 27;23(19):11388. doi: 10.3390/ijms231911388. Int J Mol Sci. 2022. PMID: 36232689 Free PMC article.
-
Antagonistic action of strigolactone and cytokinin in bud outgrowth control.Plant Physiol. 2012 Jan;158(1):487-98. doi: 10.1104/pp.111.186783. Epub 2011 Oct 31. Plant Physiol. 2012. PMID: 22042819 Free PMC article.
-
Hormonal control of shoot branching.J Exp Bot. 2008;59(1):67-74. doi: 10.1093/jxb/erm134. Epub 2007 Aug 28. J Exp Bot. 2008. PMID: 17728300 Review.
-
Auxin-cytokinin interactions in the control of shoot branching.Plant Mol Biol. 2009 Mar;69(4):429-35. doi: 10.1007/s11103-008-9416-3. Epub 2008 Oct 30. Plant Mol Biol. 2009. PMID: 18974937 Review.
Cited by
-
Exogenous application of GA3 inactively regulates axillary bud outgrowth by influencing of branching-inhibitors and bud-regulating hormones in apple (Malus domestica Borkh.).Mol Genet Genomics. 2018 Dec;293(6):1547-1563. doi: 10.1007/s00438-018-1481-y. Epub 2018 Aug 16. Mol Genet Genomics. 2018. PMID: 30116947
-
Distinct gibberellin functions during and after grapevine bud dormancy release.J Exp Bot. 2018 Mar 24;69(7):1635-1648. doi: 10.1093/jxb/ery022. J Exp Bot. 2018. PMID: 29385616 Free PMC article.
-
Regulatory frameworks involved in the floral induction, formation and developmental programming of woody horticultural plants: a case study on blueberries.Front Plant Sci. 2024 Feb 12;15:1336892. doi: 10.3389/fpls.2024.1336892. eCollection 2024. Front Plant Sci. 2024. PMID: 38410737 Free PMC article. Review.
-
Molecular Responses during Plant Grafting and Its Regulation by Auxins, Cytokinins, and Gibberellins.Biomolecules. 2019 Aug 22;9(9):397. doi: 10.3390/biom9090397. Biomolecules. 2019. PMID: 31443419 Free PMC article. Review.
-
Genome-wide analysis of TCP transcription factor family in sunflower and identification of HaTCP1 involved in the regulation of shoot branching.BMC Plant Biol. 2023 Apr 27;23(1):222. doi: 10.1186/s12870-023-04211-0. BMC Plant Biol. 2023. PMID: 37101166 Free PMC article.
References
-
- Agharkar M., Lomba P., Altpeter F., Zhang H., Kenworthy K., Lange T. (2007) Stable expression of AtGA2ox1 in a low-input turfgrass (Paspalum notatum Flugge) reduces bioactive gibberellin levels and improves turf quality under field conditions. Plant Biotechnol. J. 5: 791–801. - PubMed
-
- Bangerth F. (1994) Response of cytokinin concentration in the xylem exudate of bean (Phaseolus vulgaris L.) plants to decapitation and auxin treatment, and relationship to apical dominance. Planta 194: 439–442.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
