

FTD/ALS-associated poly(GR) protein impairs the Notch pathway and is recruited by poly(GA) into cytoplasmic inclusions
- PMID: 26031661
- PMCID: PMC4575383
- DOI: 10.1007/s00401-015-1448-6
FTD/ALS-associated poly(GR) protein impairs the Notch pathway and is recruited by poly(GA) into cytoplasmic inclusions
Abstract
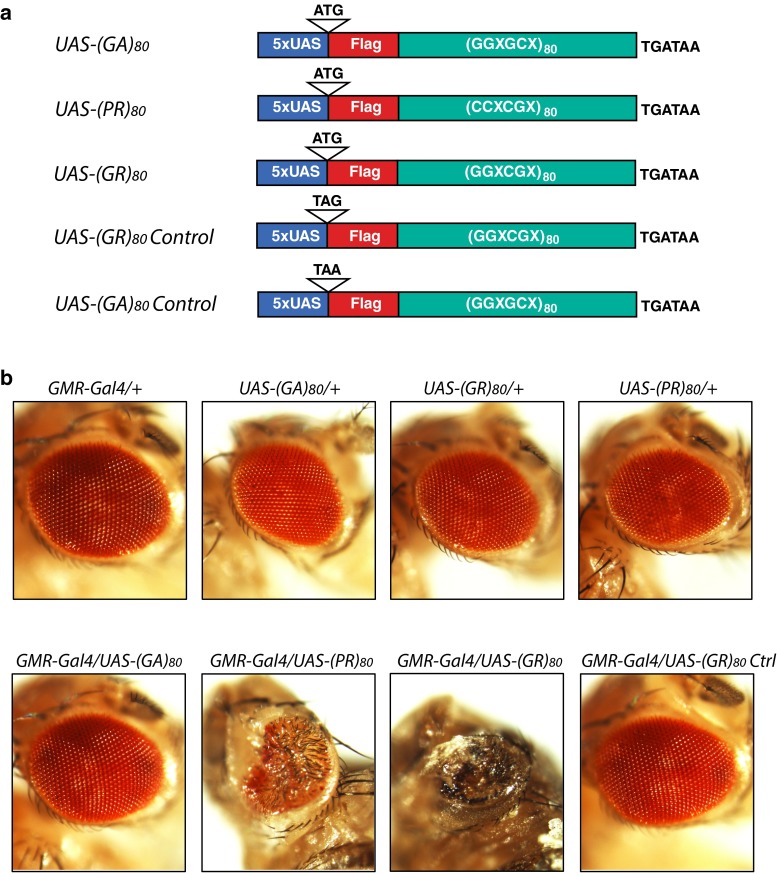
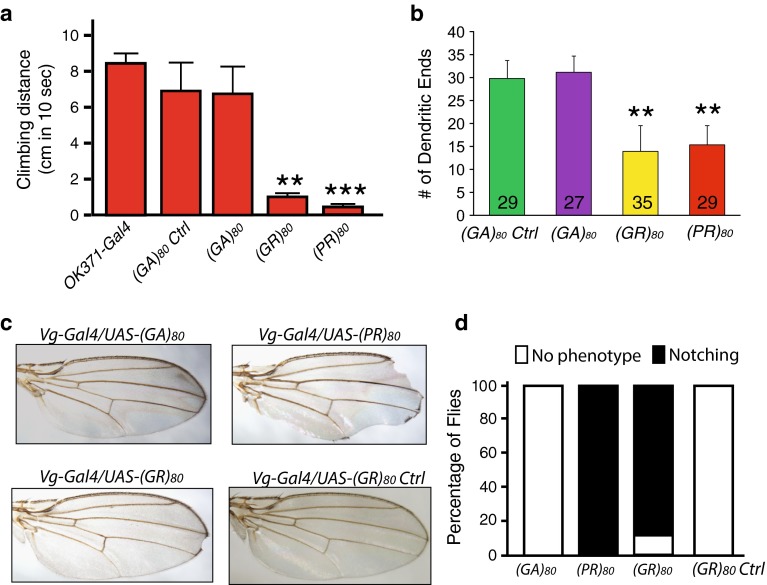
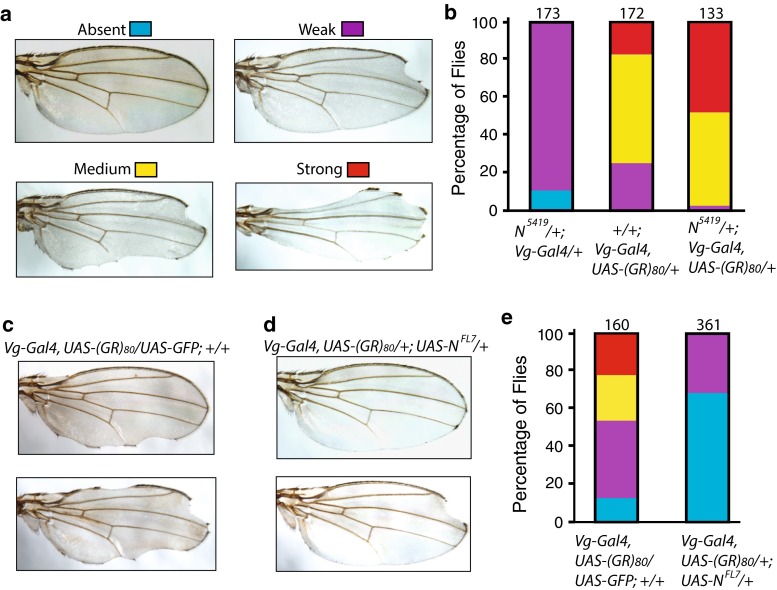
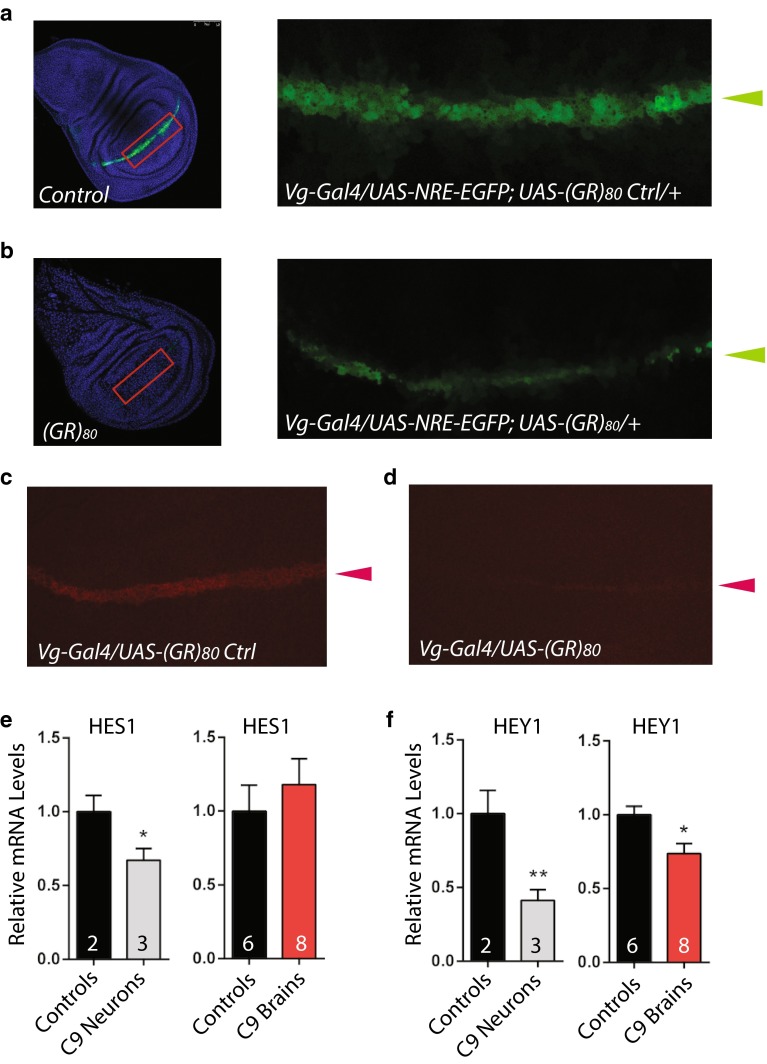
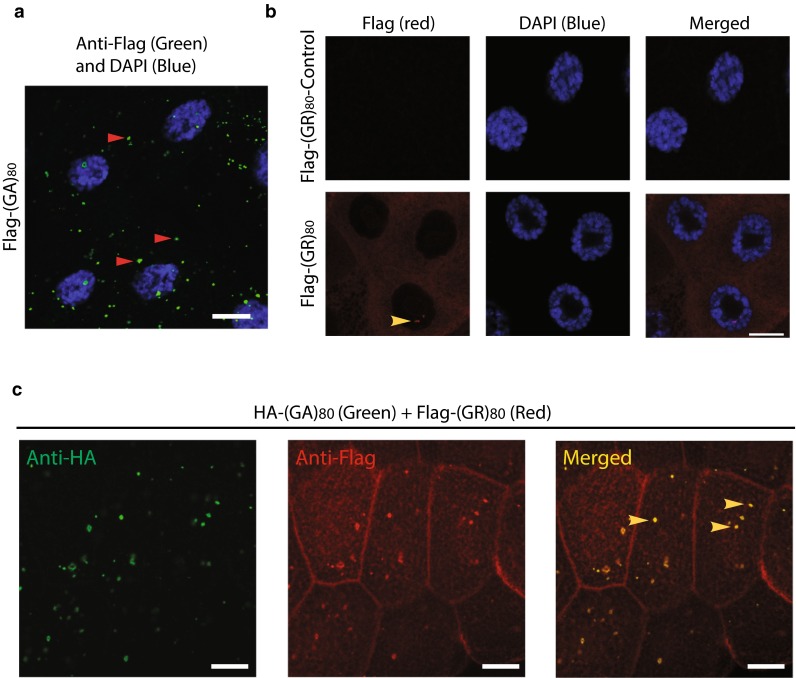
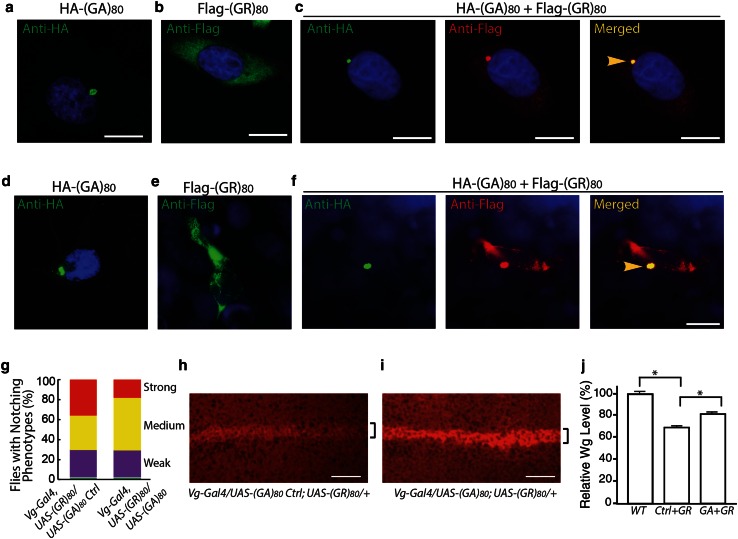
C9ORF72 repeat expansion is the most common genetic mutation in frontotemporal dementia (FTD) and amyotrophic lateral sclerosis (ALS). Abnormal dipeptide repeat proteins (DPRs) generated from repeat-associated non-AUG (RAN) translation of repeat-containing RNAs are thought to be pathogenic; however, the mechanisms are unknown. Here we report that (GR)80 and (PR)80 are toxic in neuronal and non-neuronal cells in Drosophila. In contrast to reported shorter poly(GR) forms, (GR)80 is mostly localized throughout the cytosol without detectable accumulation in the nucleolus, accompanied by suppression of Notch signaling and cell loss in the wing. Some Notch target genes are also downregulated in brains and iPSC-derived cortical neurons of C9ORF72 patients. Increased Notch expression largely suppressed (GR)80-induced cell loss in the wing. When co-expressed in Drosophila, HeLa cells, or human neurons, (GA)80 recruited (GR)80 into cytoplasmic inclusions, partially decreasing the toxicity of (GR)80 and restoring Notch signaling in Drosophila. Thus, different DPRs have opposing roles in cell loss and we identify the Notch pathway as one of the receptor signaling pathways that might be compromised in C9ORF72 FTD/ALS.
Keywords: ALS; DPR; Drosophila; FTD; Inclusion; Motor neuron; Notch; Poly(GA); Poly(GR); Poly(PR); RAN translation.
Figures
Similar articles
-
Poly(GR) in C9ORF72-Related ALS/FTD Compromises Mitochondrial Function and Increases Oxidative Stress and DNA Damage in iPSC-Derived Motor Neurons.Neuron. 2016 Oct 19;92(2):383-391. doi: 10.1016/j.neuron.2016.09.015. Epub 2016 Oct 6. Neuron. 2016. PMID: 27720481 Free PMC article.
-
Characterization of the dipeptide repeat protein in the molecular pathogenesis of c9FTD/ALS.Hum Mol Genet. 2015 Mar 15;24(6):1630-45. doi: 10.1093/hmg/ddu576. Epub 2014 Nov 14. Hum Mol Genet. 2015. PMID: 25398948
-
Distribution of dipeptide repeat proteins in cellular models and C9orf72 mutation cases suggests link to transcriptional silencing.Acta Neuropathol. 2015 Oct;130(4):537-55. doi: 10.1007/s00401-015-1450-z. Epub 2015 Jun 18. Acta Neuropathol. 2015. PMID: 26085200 Free PMC article.
-
Insights into C9ORF72-Related ALS/FTD from Drosophila and iPSC Models.Trends Neurosci. 2018 Jul;41(7):457-469. doi: 10.1016/j.tins.2018.04.002. Epub 2018 May 2. Trends Neurosci. 2018. PMID: 29729808 Free PMC article. Review.
-
Arginine-rich dipeptide-repeat proteins as phase disruptors in C9-ALS/FTD.Emerg Top Life Sci. 2020 Dec 11;4(3):293-305. doi: 10.1042/ETLS20190167. Emerg Top Life Sci. 2020. PMID: 32639008 Free PMC article. Review.
Cited by
-
The exocyst subunit EXOC2 regulates the toxicity of expanded GGGGCC repeats in C9ORF72-ALS/FTD.Cell Rep. 2024 Jul 23;43(7):114375. doi: 10.1016/j.celrep.2024.114375. Epub 2024 Jun 26. Cell Rep. 2024. PMID: 38935506 Free PMC article.
-
Poly(GR) in C9ORF72-Related ALS/FTD Compromises Mitochondrial Function and Increases Oxidative Stress and DNA Damage in iPSC-Derived Motor Neurons.Neuron. 2016 Oct 19;92(2):383-391. doi: 10.1016/j.neuron.2016.09.015. Epub 2016 Oct 6. Neuron. 2016. PMID: 27720481 Free PMC article.
-
Mice deficient in the C-terminal domain of TAR DNA-binding protein 43 develop age-dependent motor dysfunction associated with impaired Notch1-Akt signaling pathway.Acta Neuropathol Commun. 2019 Jul 25;7(1):118. doi: 10.1186/s40478-019-0776-5. Acta Neuropathol Commun. 2019. PMID: 31345270 Free PMC article.
-
Downregulation of exosomal miR-204-5p and miR-632 as a biomarker for FTD: a GENFI study.J Neurol Neurosurg Psychiatry. 2018 Aug;89(8):851-858. doi: 10.1136/jnnp-2017-317492. Epub 2018 Feb 6. J Neurol Neurosurg Psychiatry. 2018. PMID: 29434051 Free PMC article.
-
Post-Developmental Roles of Notch Signaling in the Nervous System.Biomolecules. 2020 Jul 1;10(7):985. doi: 10.3390/biom10070985. Biomolecules. 2020. PMID: 32630239 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
