

Ltc1 is an ER-localized sterol transporter and a component of ER-mitochondria and ER-vacuole contacts
- PMID: 25987606
- PMCID: PMC4442815
- DOI: 10.1083/jcb.201502033
Ltc1 is an ER-localized sterol transporter and a component of ER-mitochondria and ER-vacuole contacts
Abstract
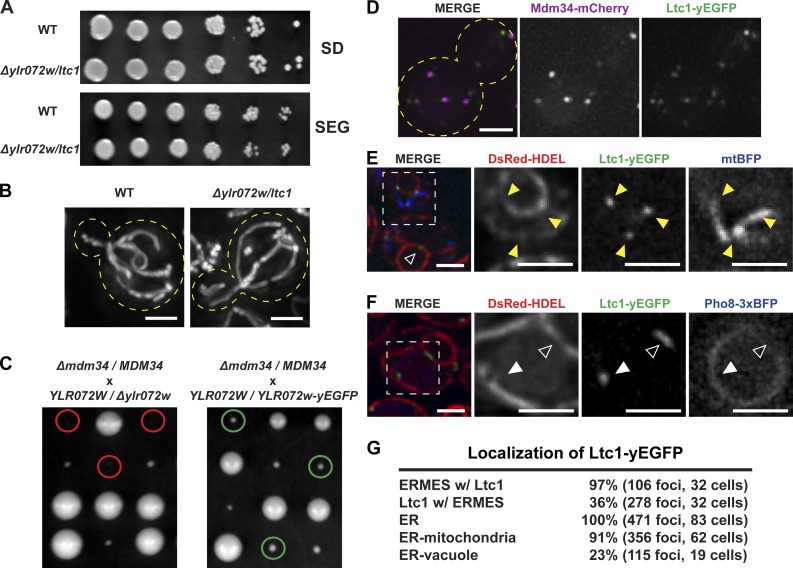
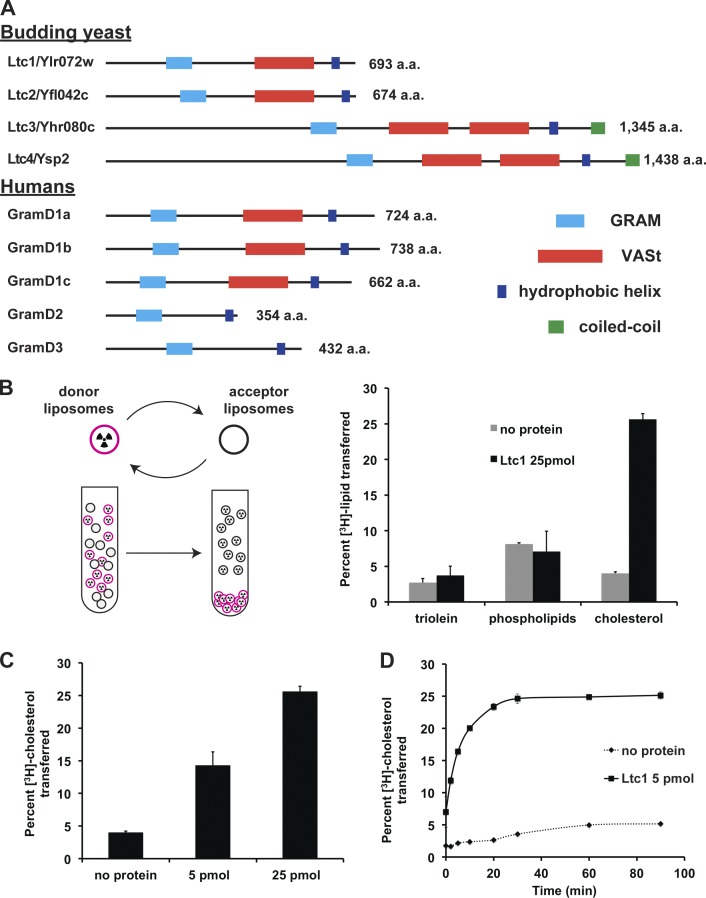
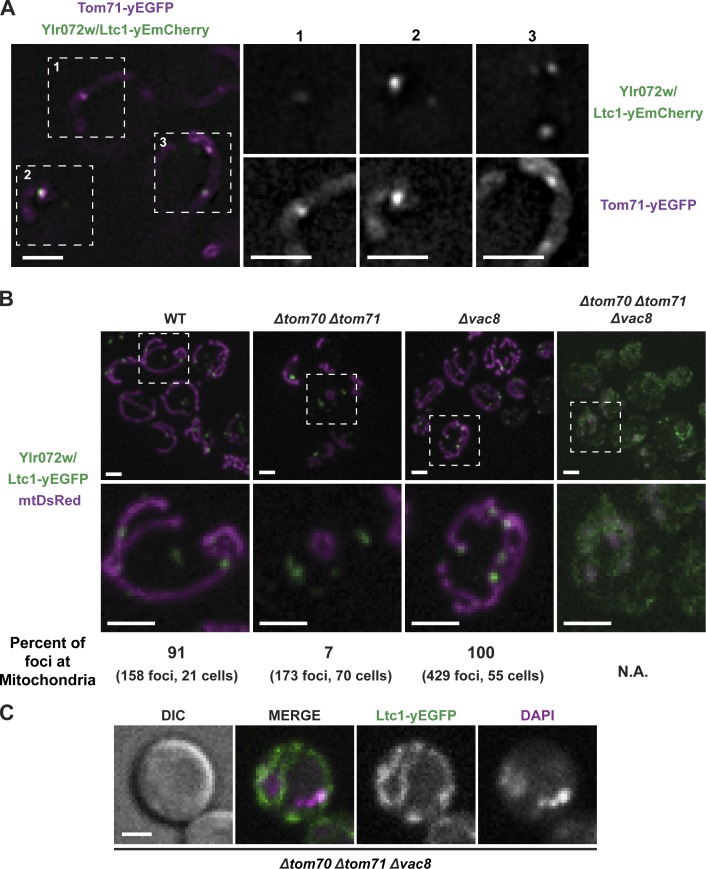
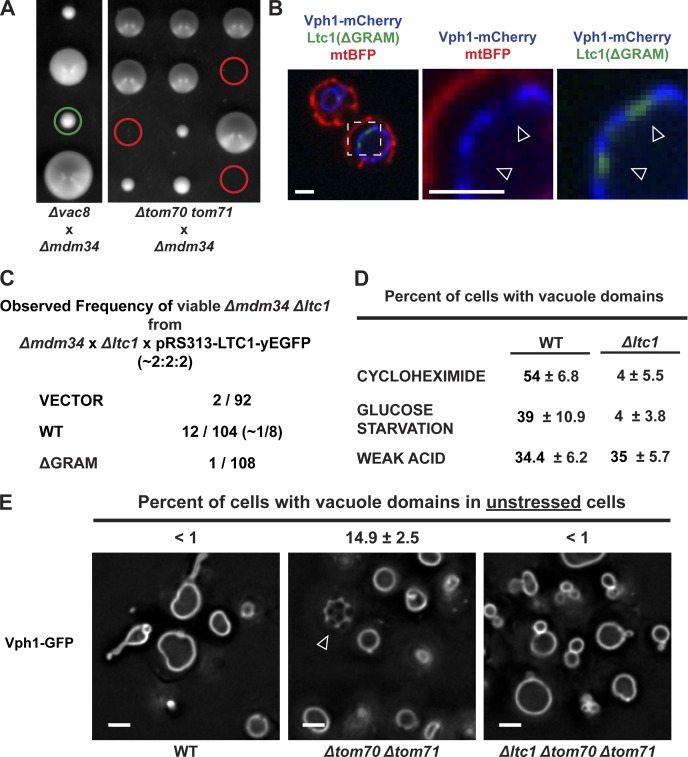
Organelle contact sites perform fundamental functions in cells, including lipid and ion homeostasis, membrane dynamics, and signaling. Using a forward proteomics approach in yeast, we identified new ER-mitochondria and ER-vacuole contacts specified by an uncharacterized protein, Ylr072w. Ylr072w is a conserved protein with GRAM and VASt domains that selectively transports sterols and is thus termed Ltc1, for Lipid transfer at contact site 1. Ltc1 localized to ER-mitochondria and ER-vacuole contacts via the mitochondrial import receptors Tom70/71 and the vacuolar protein Vac8, respectively. At mitochondria, Ltc1 was required for cell viability in the absence of Mdm34, a subunit of the ER-mitochondria encounter structure. At vacuoles, Ltc1 was required for sterol-enriched membrane domain formation in response to stress. Increasing the proportion of Ltc1 at vacuoles was sufficient to induce sterol-enriched vacuolar domains without stress. Thus, our data support a model in which Ltc1 is a sterol-dependent regulator of organelle and cellular homeostasis via its dual localization to ER-mitochondria and ER-vacuole contact sites.
© 2015 Murley et al.
Figures
Similar articles
-
Vps13-Mcp1 interact at vacuole-mitochondria interfaces and bypass ER-mitochondria contact sites.J Cell Biol. 2017 Oct 2;216(10):3219-3229. doi: 10.1083/jcb.201610055. Epub 2017 Sep 1. J Cell Biol. 2017. PMID: 28864540 Free PMC article.
-
Lam6 Regulates the Extent of Contacts between Organelles.Cell Rep. 2015 Jul 7;12(1):7-14. doi: 10.1016/j.celrep.2015.06.022. Epub 2015 Jun 25. Cell Rep. 2015. PMID: 26119743 Free PMC article.
-
Cellular metabolism regulates contact sites between vacuoles and mitochondria.Dev Cell. 2014 Jul 14;30(1):86-94. doi: 10.1016/j.devcel.2014.06.006. Dev Cell. 2014. PMID: 25026035
-
A close-up view of membrane contact sites between the endoplasmic reticulum and the endolysosomal system: from yeast to man.Crit Rev Biochem Mol Biol. 2014 May-Jun;49(3):262-8. doi: 10.3109/10409238.2013.875512. Epub 2014 Jan 2. Crit Rev Biochem Mol Biol. 2014. PMID: 24382115 Review.
-
Organelle contact zones as sites for lipid transfer.J Biochem. 2019 Feb 1;165(2):115-123. doi: 10.1093/jb/mvy088. J Biochem. 2019. PMID: 30371789 Review.
Cited by
-
Mitochondria: multifaceted regulators of aging.BMB Rep. 2019 Jan;52(1):13-23. doi: 10.5483/BMBRep.2019.52.1.300. BMB Rep. 2019. PMID: 30545443 Free PMC article. Review.
-
The Expanding and Unexpected Functions of Mitochondria Contact Sites.Trends Cell Biol. 2019 Jul;29(7):580-590. doi: 10.1016/j.tcb.2019.02.009. Epub 2019 Mar 28. Trends Cell Biol. 2019. PMID: 30929794 Free PMC article. Review.
-
The UbiB family member Cqd1 forms a novel membrane contact site in mitochondria.J Cell Sci. 2023 May 15;136(10):jcs260578. doi: 10.1242/jcs.260578. Epub 2023 May 18. J Cell Sci. 2023. PMID: 37073556 Free PMC article.
-
A Systematic Protein Turnover Map for Decoding Protein Degradation.Cell Rep. 2020 Nov 10;33(6):108378. doi: 10.1016/j.celrep.2020.108378. Cell Rep. 2020. PMID: 33176155 Free PMC article.
-
Nutrient Signaling, Stress Response, and Inter-organelle Communication Are Non-canonical Determinants of Cell Fate.Cell Rep. 2020 Dec 1;33(9):108446. doi: 10.1016/j.celrep.2020.108446. Cell Rep. 2020. PMID: 33264609 Free PMC article.
References
-
- Bartlett G.R. 1959. Colorimetric assay methods for free and phosphorylated glyceric acids. J. Biol. Chem. 234:469–471. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
