

Live-cell imaging of actin dynamics reveals mechanisms of stereocilia length regulation in the inner ear
- PMID: 25898120
- PMCID: PMC4411292
- DOI: 10.1038/ncomms7873
Live-cell imaging of actin dynamics reveals mechanisms of stereocilia length regulation in the inner ear
Abstract
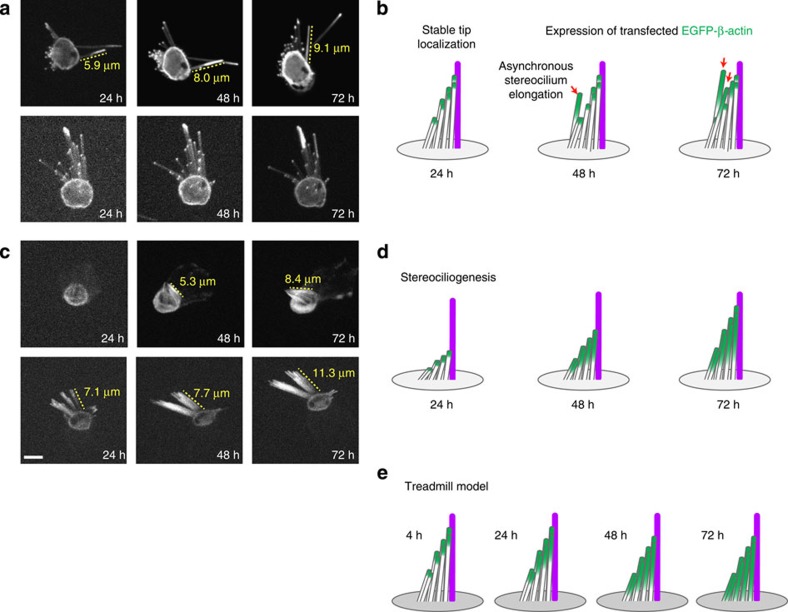
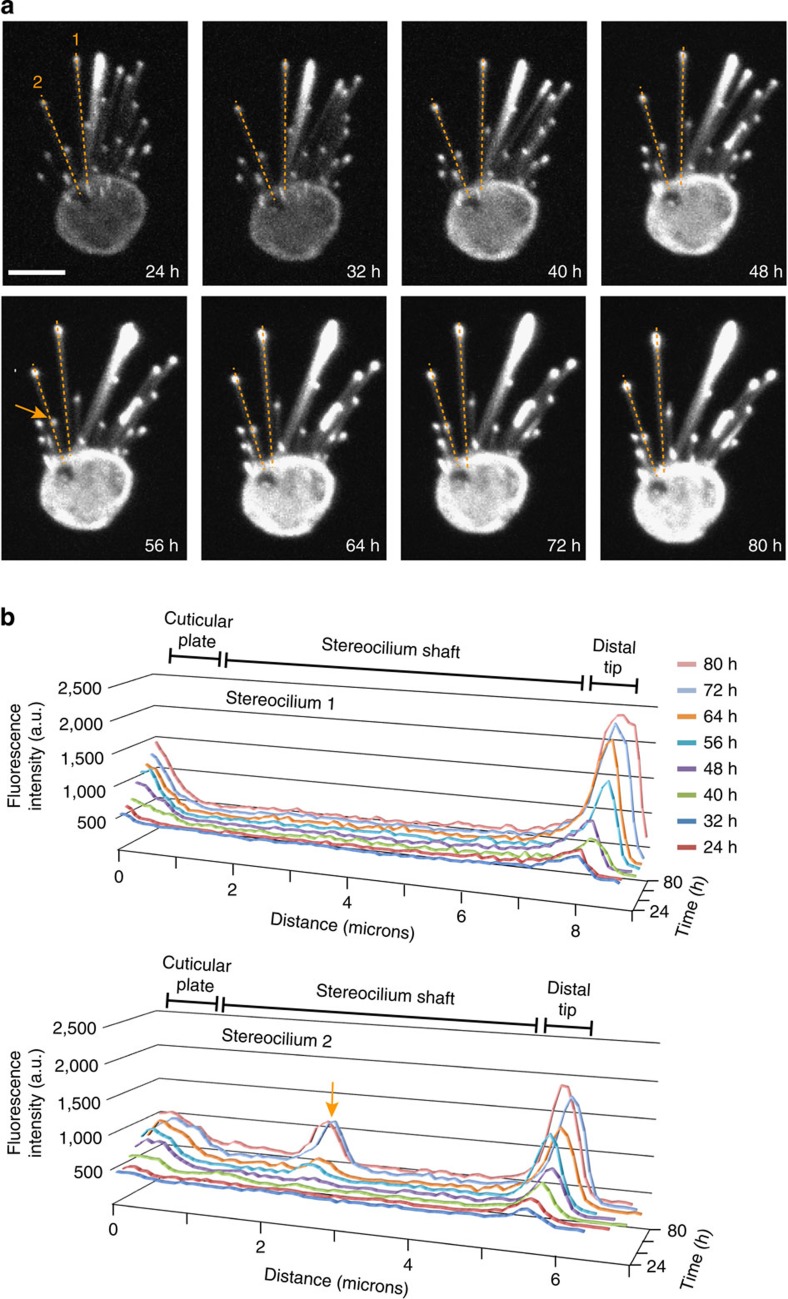
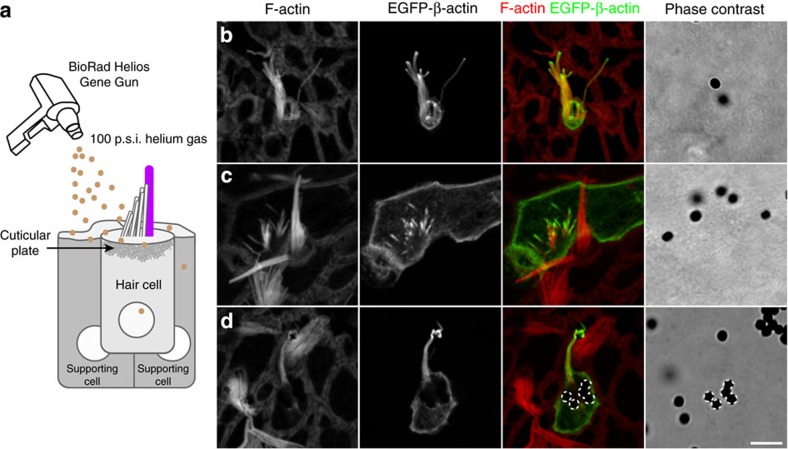
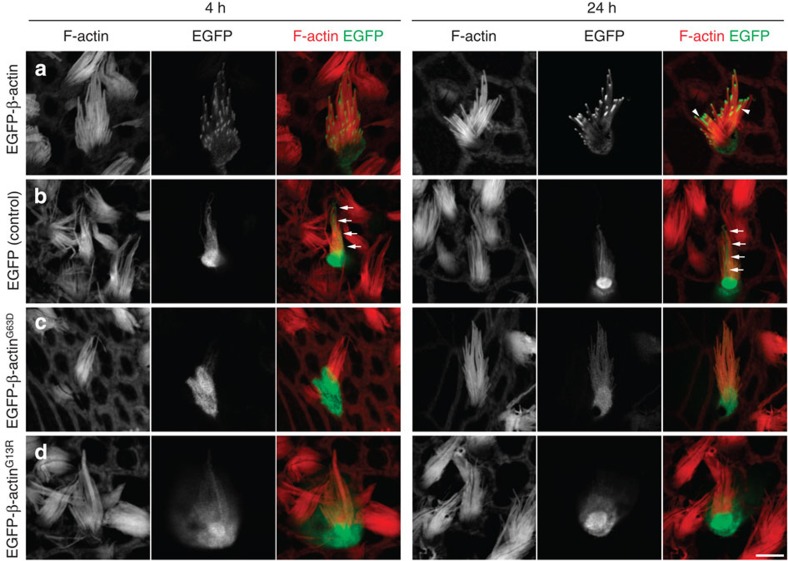
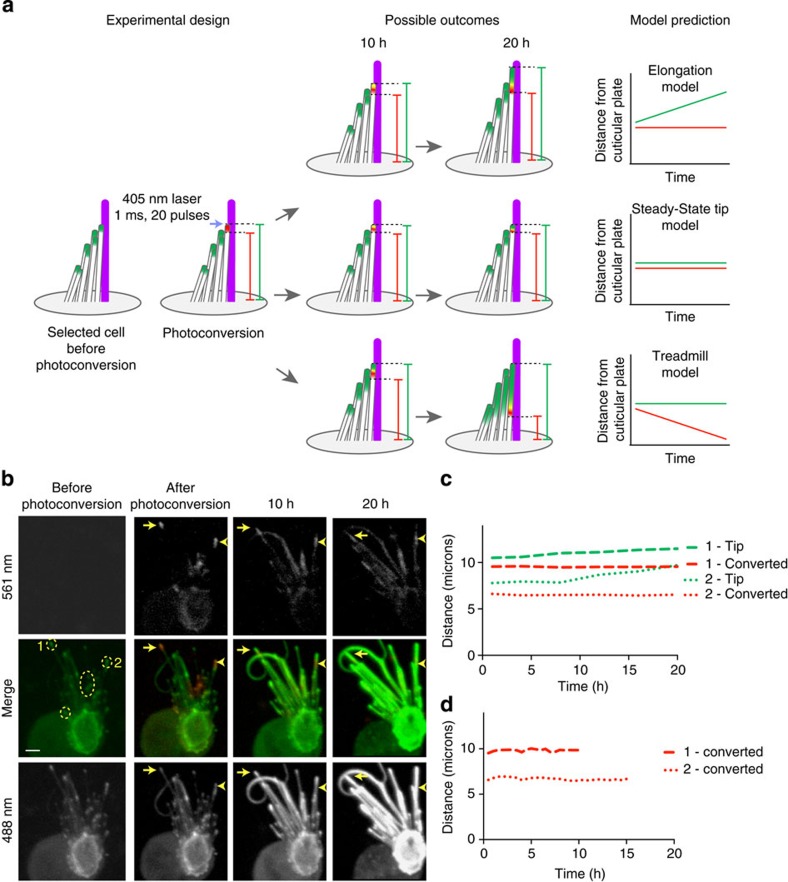
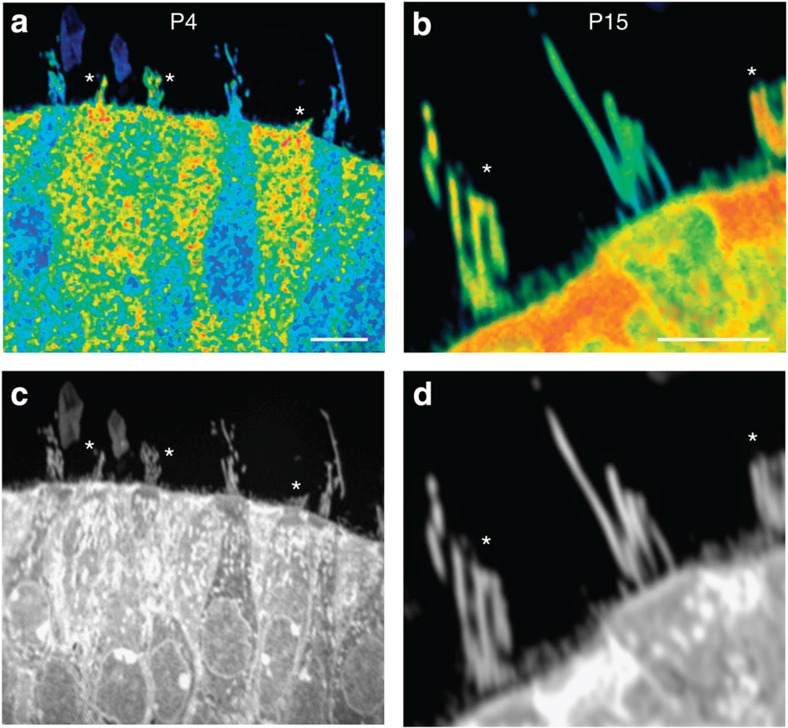
The maintenance of sensory hair cell stereocilia is critical for lifelong hearing; however, mechanisms of structural homeostasis remain poorly understood. Conflicting models propose that stereocilia F-actin cores are either continually renewed every 24-48 h via a treadmill or are stable, exceptionally long-lived structures. Here to distinguish between these models, we perform an unbiased survey of stereocilia actin dynamics in more than 500 utricle hair cells. Live-imaging EGFP-β-actin or dendra2-β-actin reveal stable F-actin cores with turnover and elongation restricted to stereocilia tips. Fixed-cell microscopy of wild-type and mutant β-actin demonstrates that incorporation of actin monomers into filaments is required for localization to stereocilia tips. Multi-isotope imaging mass spectrometry and live imaging of single differentiating hair cells capture stereociliogenesis and explain uniform incorporation of (15)N-labelled protein and EGFP-β-actin into nascent stereocilia. Collectively, our analyses support a model in which stereocilia actin cores are stable structures that incorporate new F-actin only at the distal tips.
Figures
Similar articles
-
Stereocilia morphogenesis and maintenance through regulation of actin stability.Semin Cell Dev Biol. 2017 May;65:88-95. doi: 10.1016/j.semcdb.2016.08.017. Epub 2016 Aug 23. Semin Cell Dev Biol. 2017. PMID: 27565685 Free PMC article. Review.
-
Multi-isotope imaging mass spectrometry reveals slow protein turnover in hair-cell stereocilia.Nature. 2012 Jan 15;481(7382):520-4. doi: 10.1038/nature10745. Nature. 2012. PMID: 22246323 Free PMC article.
-
The stable actin core of mechanosensory stereocilia features continuous turnover of actin cross-linkers.Mol Biol Cell. 2018 Aug 1;29(15):1856-1865. doi: 10.1091/mbc.E18-03-0196. Epub 2018 Jun 6. Mol Biol Cell. 2018. PMID: 29874122 Free PMC article.
-
Length regulation of mechanosensitive stereocilia depends on very slow actin dynamics and filament-severing proteins.Nat Commun. 2015 Apr 21;6:6855. doi: 10.1038/ncomms7855. Nat Commun. 2015. PMID: 25897778 Free PMC article.
-
Human deafness-associated variants alter the dynamics of key molecules in hair cell stereocilia F-actin cores.Hum Genet. 2022 Apr;141(3-4):363-382. doi: 10.1007/s00439-021-02304-0. Epub 2021 Jul 7. Hum Genet. 2022. PMID: 34232383 Free PMC article. Review.
Cited by
-
The Rho GTPase Cell Division Cycle 42 Regulates Stereocilia Development in Cochlear Hair Cells.Front Cell Dev Biol. 2021 Oct 22;9:765559. doi: 10.3389/fcell.2021.765559. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34746154 Free PMC article.
-
Myosin-XVa Controls Both Staircase Architecture and Diameter Gradation of Stereocilia Rows in the Auditory Hair Cell Bundles.J Assoc Res Otolaryngol. 2020 Apr;21(2):121-135. doi: 10.1007/s10162-020-00745-4. Epub 2020 Mar 9. J Assoc Res Otolaryngol. 2020. PMID: 32152769 Free PMC article.
-
Stereocilia morphogenesis and maintenance through regulation of actin stability.Semin Cell Dev Biol. 2017 May;65:88-95. doi: 10.1016/j.semcdb.2016.08.017. Epub 2016 Aug 23. Semin Cell Dev Biol. 2017. PMID: 27565685 Free PMC article. Review.
-
Cy3-ATP labeling of unfixed, permeabilized mouse hair cells.Sci Rep. 2021 Dec 13;11(1):23855. doi: 10.1038/s41598-021-03365-x. Sci Rep. 2021. PMID: 34903829 Free PMC article.
-
The Stereociliary Paracrystal Is a Dynamic Cytoskeletal Scaffold In Vivo.Cell Rep. 2015 Nov 17;13(7):1287-1294. doi: 10.1016/j.celrep.2015.10.003. Epub 2015 Nov 5. Cell Rep. 2015. PMID: 26549442 Free PMC article.
References
-
- Nayak G. D., Ratnayaka H. S., Goodyear R. J. & Richardson G. P. Development of the hair bundle and mechanotransduction. Int. J. Dev. Biol. 51, 597–608 (2007). - PubMed
-
- Tilney L. G., Tilney M. S. & DeRosier D. J. Actin filaments, stereocilia, and hair cells: how cells count and measure. Annu. Rev. Cell Biol. 8, 257–274 (1992). - PubMed
-
- Kelley M. W. Cellular commitment and differentiation in the organ of Corti. Int. J. Dev. Biol. 51, 571–583 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01DC03463/DC/NIDCD NIH HHS/United States
- P41EB001974/EB/NIBIB NIH HHS/United States
- R01 DC004179/DC/NIDCD NIH HHS/United States
- R37DK39773/DK/NIDDK NIH HHS/United States
- DC000079-02/DC/NIDCD NIH HHS/United States
- R01DC04179/DC/NIDCD NIH HHS/United States
- P41RR14579/RR/NCRR NIH HHS/United States
- T32 DC000039/DC/NIDCD NIH HHS/United States
- P41 EB001974/EB/NIBIB NIH HHS/United States
- Z01 DC000039/ImNIH/Intramural NIH HHS/United States
- R01GM47214/GM/NIGMS NIH HHS/United States
- R01 EY012963/EY/NEI NIH HHS/United States
- DC000039-17/DC/NIDCD NIH HHS/United States
- R01 DC002281/DC/NIDCD NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R01 GM047214/GM/NIGMS NIH HHS/United States
- R01DC00033/DC/NIDCD NIH HHS/United States
- R01EY12963/EY/NEI NIH HHS/United States
- R37 DK039773/DK/NIDDK NIH HHS/United States
- Z01 DC000033/ImNIH/Intramural NIH HHS/United States
- R01D K58762/PHS HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
