

HIV Blocks Interferon Induction in Human Dendritic Cells and Macrophages by Dysregulation of TBK1
- PMID: 25855743
- PMCID: PMC4468486
- DOI: 10.1128/JVI.00889-15
HIV Blocks Interferon Induction in Human Dendritic Cells and Macrophages by Dysregulation of TBK1
Abstract
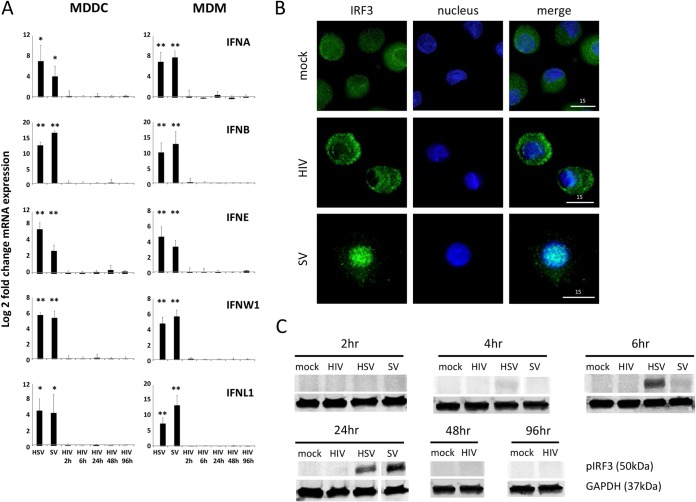
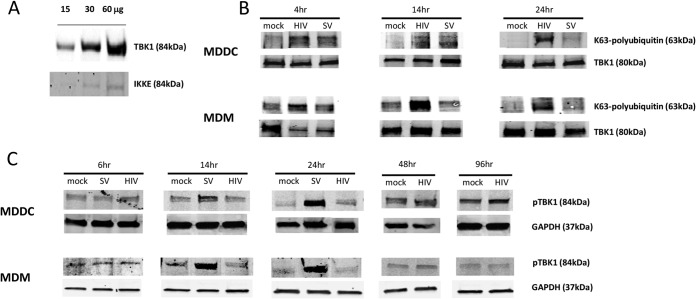
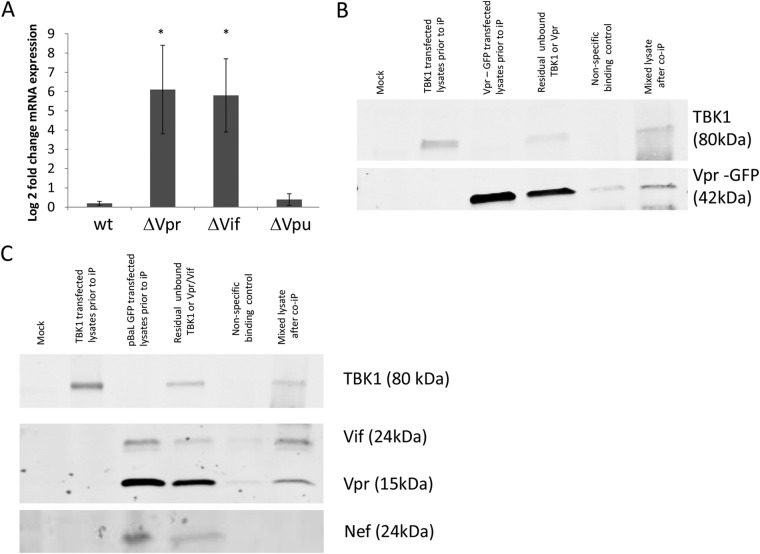
Dendritic cells (DCs) and macrophages are present in the tissues of the anogenital tract, where HIV-1 transmission occurs in almost all cases. These cells are both target cells for HIV-1 and represent the first opportunity for the virus to interfere with innate recognition. Previously we have shown that both cell types fail to produce type I interferons (IFNs) in response to HIV-1 but that, unlike T cells, the virus does not block IFN induction by targeting IFN regulatory factor 3 (IRF3) for cellular degradation. Thus, either HIV-1 inhibits IFN induction by an alternate mechanism or, less likely, these cells fail to sense HIV-1. Here we show that HIV-1 (but not herpes simplex virus 2 [HSV-2] or Sendai virus)-exposed DCs and macrophages fail to induce the expression of all known type I and III IFN genes. These cells do sense the virus, and pattern recognition receptor (PRR)-induced signaling pathways are triggered. The precise stage in the IFN-inducing signaling pathway that HIV-1 targets to block IFN induction was identified; phosphorylation but not K63 polyubiquitination of TANK-binding kinase 1 (TBK1) was completely inhibited. Two HIV-1 accessory proteins, Vpr and Vif, were shown to bind to TBK1, and their individual deletion partly restored IFN-β expression. Thus, the inhibition of TBK1 autophosphorylation by binding of these proteins appears to be the principal mechanism by which HIV-1 blocks type I and III IFN induction in myeloid cells.
Importance: Dendritic cells (DCs) and macrophages are key HIV target cells. Therefore, definition of how HIV impairs innate immune responses to initially establish infection is essential to design preventative interventions, especially by restoring initial interferon production. Here we demonstrate how HIV-1 blocks interferon induction by inhibiting the function of a key kinase in the interferon signaling pathway, TBK1, via two different viral accessory proteins. Other viral proteins have been shown to target the general effects of TBK1, but this precise targeting between ubiquitination and phosphorylation of TBK1 is novel.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures


Similar articles
-
Dengue Virus NS Proteins Inhibit RIG-I/MAVS Signaling by Blocking TBK1/IRF3 Phosphorylation: Dengue Virus Serotype 1 NS4A Is a Unique Interferon-Regulating Virulence Determinant.mBio. 2015 May 12;6(3):e00553-15. doi: 10.1128/mBio.00553-15. mBio. 2015. PMID: 25968648 Free PMC article.
-
Role of Herpes Simplex Virus 1 γ34.5 in the Regulation of IRF3 Signaling.J Virol. 2017 Nov 14;91(23):e01156-17. doi: 10.1128/JVI.01156-17. Print 2017 Dec 1. J Virol. 2017. PMID: 28904192 Free PMC article.
-
Herpes Simplex Virus 1 Serine Protease VP24 Blocks the DNA-Sensing Signal Pathway by Abrogating Activation of Interferon Regulatory Factor 3.J Virol. 2016 May 27;90(12):5824-5829. doi: 10.1128/JVI.00186-16. Print 2016 Jun 15. J Virol. 2016. PMID: 27076640 Free PMC article.
-
Identification and utility of innate immune system evasion mechanisms of ASFV.Virus Res. 2013 Apr;173(1):87-100. doi: 10.1016/j.virusres.2012.10.013. Epub 2012 Nov 16. Virus Res. 2013. PMID: 23165138 Review.
-
HIV-1, interferon and the interferon regulatory factor system: an interplay between induction, antiviral responses and viral evasion.Cytokine Growth Factor Rev. 2012 Aug-Oct;23(4-5):255-70. doi: 10.1016/j.cytogfr.2012.06.001. Epub 2012 Jun 27. Cytokine Growth Factor Rev. 2012. PMID: 22748237 Review.
Cited by
-
HIV-induced RSAD2/Viperin supports sustained infection of monocyte-derived macrophages.J Virol. 2024 Oct 22;98(10):e0086324. doi: 10.1128/jvi.00863-24. Epub 2024 Sep 11. J Virol. 2024. PMID: 39258908
-
Lnc-RPS6P3 Inhibits Influenza A Virus Replication and Attenuates the Inhibitory Effect of NS1 on Innate Immune Response.Microorganisms. 2024 Mar 26;12(4):654. doi: 10.3390/microorganisms12040654. Microorganisms. 2024. PMID: 38674599 Free PMC article.
-
Dynamic modulation of the non-canonical NF-κB signaling pathway for HIV shock and kill.Front Cell Infect Microbiol. 2024 Mar 5;14:1354502. doi: 10.3389/fcimb.2024.1354502. eCollection 2024. Front Cell Infect Microbiol. 2024. PMID: 38505285 Free PMC article. Review.
-
Immunomodulatory Role of Interferons in Viral and Bacterial Infections.Int J Mol Sci. 2023 Jun 14;24(12):10115. doi: 10.3390/ijms241210115. Int J Mol Sci. 2023. PMID: 37373262 Free PMC article. Review.
-
HIV-1 Vpr-induced DNA damage activates NF-κB through ATM-NEMO independent of cell cycle arrest.bioRxiv [Preprint]. 2024 Jan 25:2023.05.23.541990. doi: 10.1101/2023.05.23.541990. bioRxiv. 2024. Update in: mBio. 2024 Oct 16;15(10):e0024024. doi: 10.1128/mbio.00240-24. PMID: 37292767 Free PMC article. Updated. Preprint.
References
-
- Nasr N, Lai J, Botting RA, Mercier SK, Harman AN, Kim M, Turville S, Center RJ, Domagala T, Gorry PR, Olbourne N, Cunningham AL. 2014. Inhibition of two temporal phases of HIV-1 transfer from primary Langerhans cells to T cells: the role of langerin. J Immunol 193:2554–2564. doi:10.4049/jimmunol.1400630. - DOI - PubMed
-
- Ganor Y, Zhou Z, Tudor D, Schmitt A, Vacher-Lavenu MC, Gibault L, Thiounn N, Tomasini J, Wolf JP, Bomsel M. 2010. Within 1 h, HIV-1 uses viral synapses to enter efficiently the inner, but not outer, foreskin mucosa and engages Langerhans-T cell conjugates. Mucosal Immunol 3:506–522. doi:10.1038/mi.2010.32. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
