

Positive role of promyelocytic leukemia protein in type I interferon response and its regulation by human cytomegalovirus
- PMID: 25812002
- PMCID: PMC4374831
- DOI: 10.1371/journal.ppat.1004785
Positive role of promyelocytic leukemia protein in type I interferon response and its regulation by human cytomegalovirus
Abstract
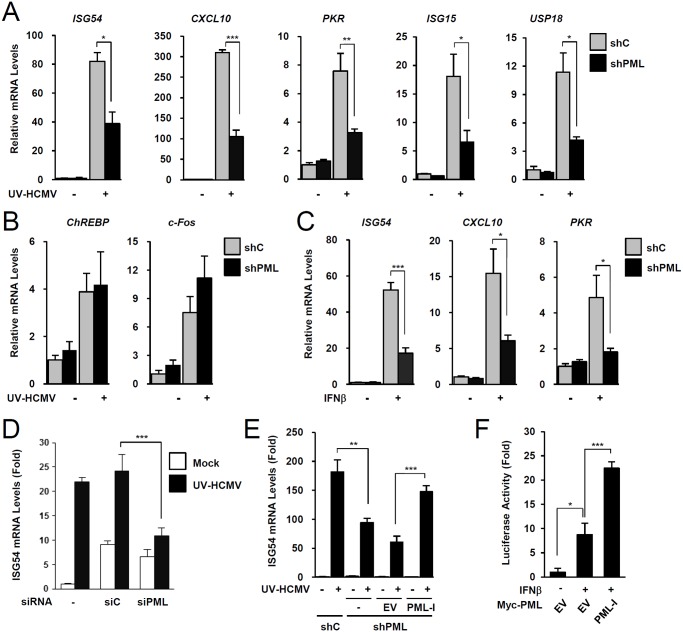
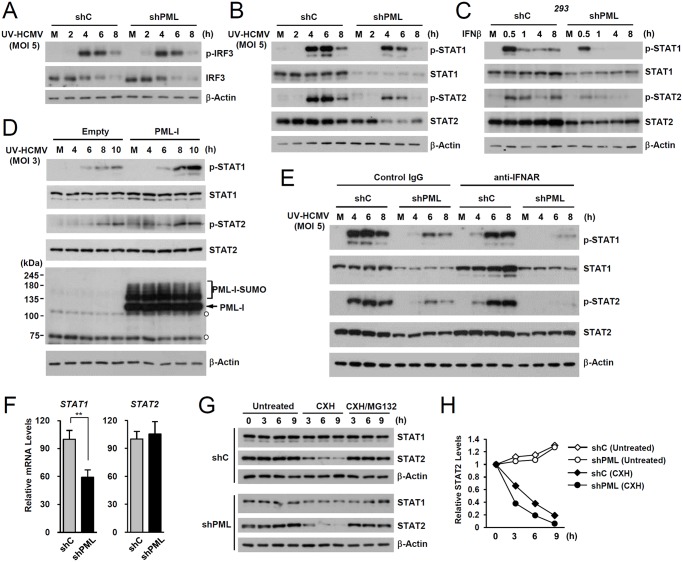
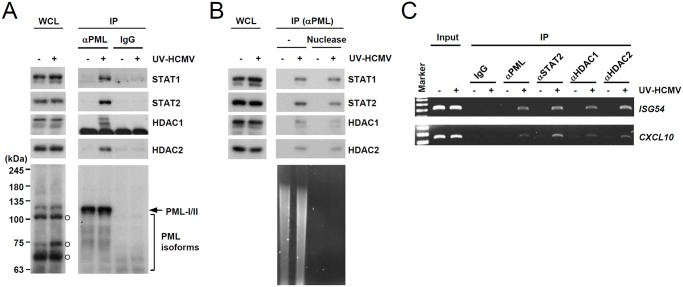
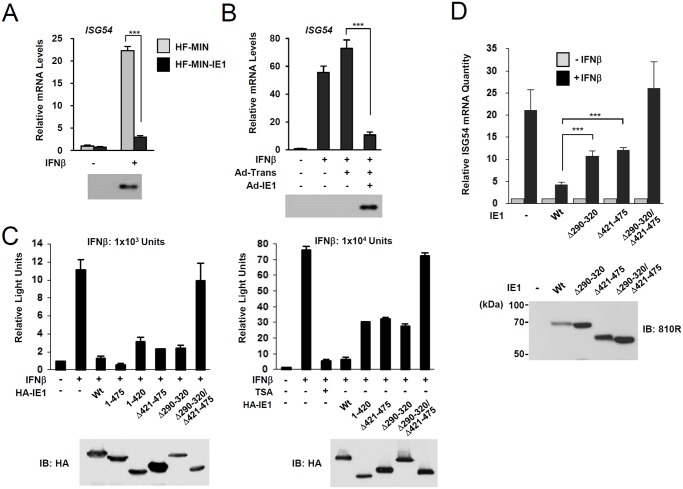
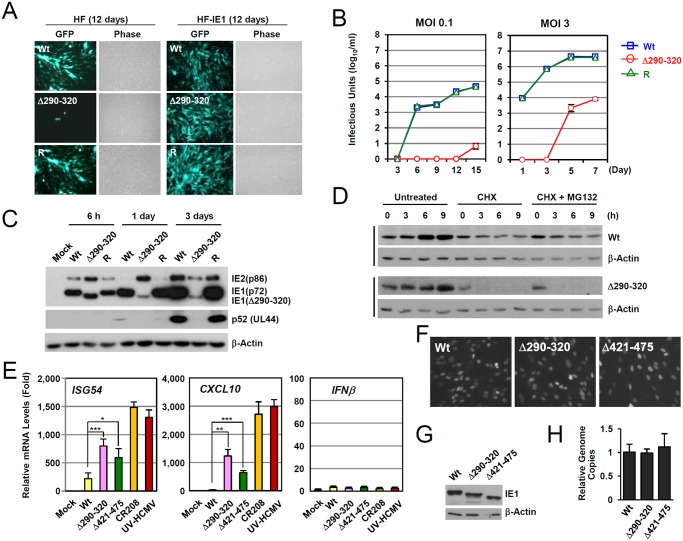
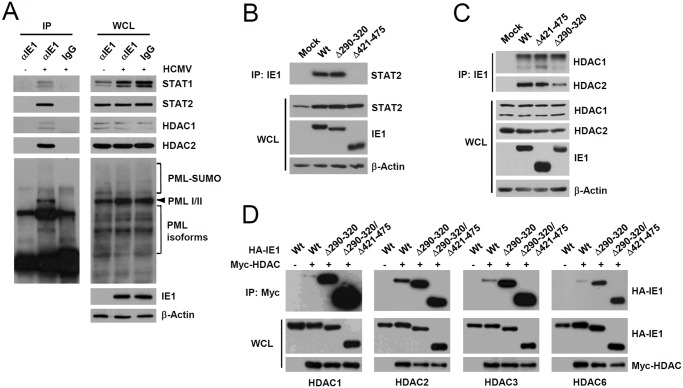
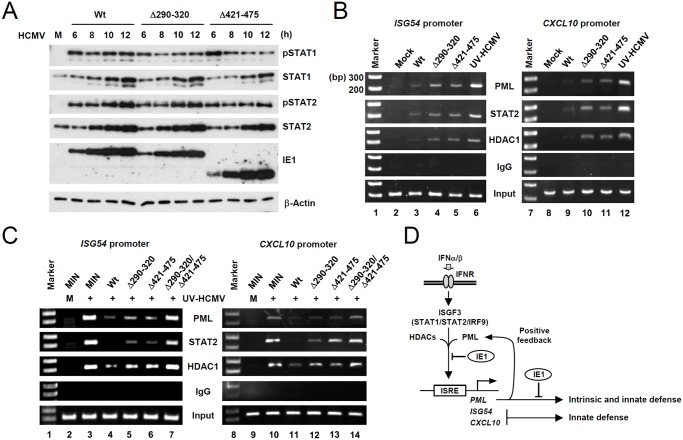
Promyelocytic leukemia protein (PML), a major component of PML nuclear bodies (also known as nuclear domain 10), is involved in diverse cellular processes such as cell proliferation, apoptosis, gene regulation, and DNA damage response. PML also acts as a restriction factor that suppresses incoming viral genomes, therefore playing an important role in intrinsic defense. Here, we show that PML positively regulates type I interferon response by promoting transcription of interferon-stimulated genes (ISGs) and that this regulation by PML is counteracted by human cytomegalovirus (HCMV) IE1 protein. Small hairpin RNA-mediated PML knockdown in human fibroblasts reduced ISG induction by treatment of interferon-β or infection with UV-inactivated HCMV. PML was required for accumulation of activated STAT1 and STAT2, interacted with them and HDAC1 and HDAC2, and was associated with ISG promoters after HCMV infection. During HCMV infection, viral IE1 protein interacted with PML, STAT1, STAT2, and HDACs. Analysis of IE1 mutant viruses revealed that, in addition to the STAT2-binding domain, the PML-binding domain of IE1 was necessary for suppression of interferon-β-mediated ISG transcription, and that IE1 inhibited ISG transcription by sequestering interferon-stimulated gene factor 3 (ISGF3) in a manner requiring its binding of PML and STAT2, but not of HDACs. In conclusion, our results demonstrate that PML participates in type I interferon-induced ISG expression by regulating ISGF3, and that this regulation by PML is counteracted by HCMV IE1, highlighting a widely shared viral strategy targeting PML to evade intrinsic and innate defense mechanisms.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Characterization of Recombinant Human Cytomegaloviruses Encoding IE1 Mutants L174P and 1-382 Reveals that Viral Targeting of PML Bodies Perturbs both Intrinsic and Innate Immune Responses.J Virol. 2015 Nov 11;90(3):1190-205. doi: 10.1128/JVI.01973-15. Print 2016 Feb 1. J Virol. 2015. PMID: 26559840 Free PMC article.
-
Disruption of PML-associated nuclear bodies by IE1 correlates with efficient early stages of viral gene expression and DNA replication in human cytomegalovirus infection.Virology. 2000 Aug 15;274(1):39-55. doi: 10.1006/viro.2000.0448. Virology. 2000. PMID: 10936087
-
Differential relocation and stability of PML-body components during productive human cytomegalovirus infection: detailed characterization by live-cell imaging.Eur J Cell Biol. 2010 Oct;89(10):757-68. doi: 10.1016/j.ejcb.2010.05.006. Eur J Cell Biol. 2010. PMID: 20599291
-
The Human CMV IE1 Protein: An Offender of PML Nuclear Bodies.Adv Anat Embryol Cell Biol. 2017;223:77-94. doi: 10.1007/978-3-319-53168-7_4. Adv Anat Embryol Cell Biol. 2017. PMID: 28528440 Review.
-
The unique role of STAT2 in constitutive and IFN-induced transcription and antiviral responses.Cytokine Growth Factor Rev. 2016 Jun;29:71-81. doi: 10.1016/j.cytogfr.2016.02.010. Epub 2016 Mar 18. Cytokine Growth Factor Rev. 2016. PMID: 27053489 Review.
Cited by
-
Crosstalk Between SUMO and Ubiquitin-Like Proteins: Implication for Antiviral Defense.Front Cell Dev Biol. 2021 Apr 21;9:671067. doi: 10.3389/fcell.2021.671067. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33968942 Free PMC article. Review.
-
Old dog, new trick: Trivalent arsenic as an immunomodulatory drug.Br J Pharmacol. 2020 May;177(10):2199-2214. doi: 10.1111/bph.15011. Epub 2020 Mar 12. Br J Pharmacol. 2020. PMID: 32022256 Free PMC article. Review.
-
Distinct temporal roles for the promyelocytic leukaemia (PML) protein in the sequential regulation of intracellular host immunity to HSV-1 infection.PLoS Pathog. 2018 Jan 8;14(1):e1006769. doi: 10.1371/journal.ppat.1006769. eCollection 2018 Jan. PLoS Pathog. 2018. PMID: 29309427 Free PMC article.
-
PML: Regulation and multifaceted function beyond tumor suppression.Cell Biosci. 2018 Jan 25;8:5. doi: 10.1186/s13578-018-0204-8. eCollection 2018. Cell Biosci. 2018. PMID: 29416846 Free PMC article. Review.
-
Swine Promyelocytic Leukemia Isoform II Inhibits Pseudorabies Virus Infection by Suppressing Viral Gene Transcription in Promyelocytic Leukemia Nuclear Bodies.J Virol. 2020 Aug 31;94(18):e01197-20. doi: 10.1128/JVI.01197-20. Print 2020 Aug 31. J Virol. 2020. PMID: 32641476 Free PMC article.
References
-
- Bhattacharya S, Eckner R, Grossman S, Oldread E, Arany Z, D'Andrea A, et al. Cooperation of Stat2 and p300/CBP in signalling induced by interferon-alpha. Nature. 1996;383(6598):344–7. - PubMed
-
- Paulson M, Press C, Smith E, Tanese N, Levy DE. IFN-Stimulated transcription through a TBP-free acetyltransferase complex escapes viral shutoff. Nat Cell Biol. 2002;4(2):140–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
