

Cytomegalovirus infection drives adaptive epigenetic diversification of NK cells with altered signaling and effector function
- PMID: 25786176
- PMCID: PMC4612277
- DOI: 10.1016/j.immuni.2015.02.008
Cytomegalovirus infection drives adaptive epigenetic diversification of NK cells with altered signaling and effector function
Abstract
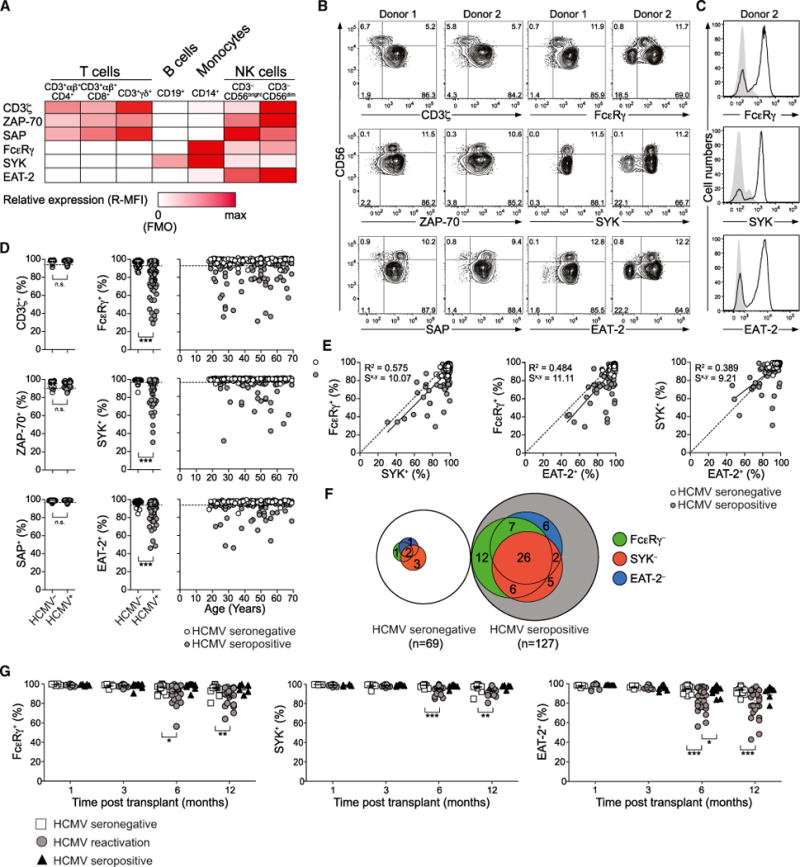
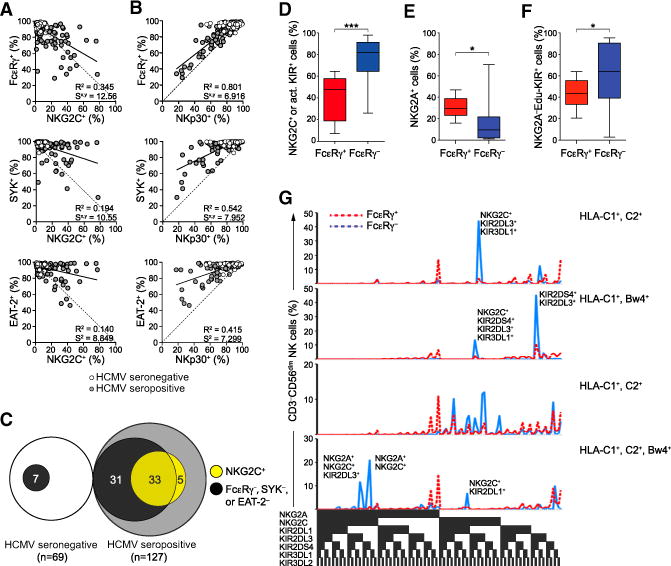
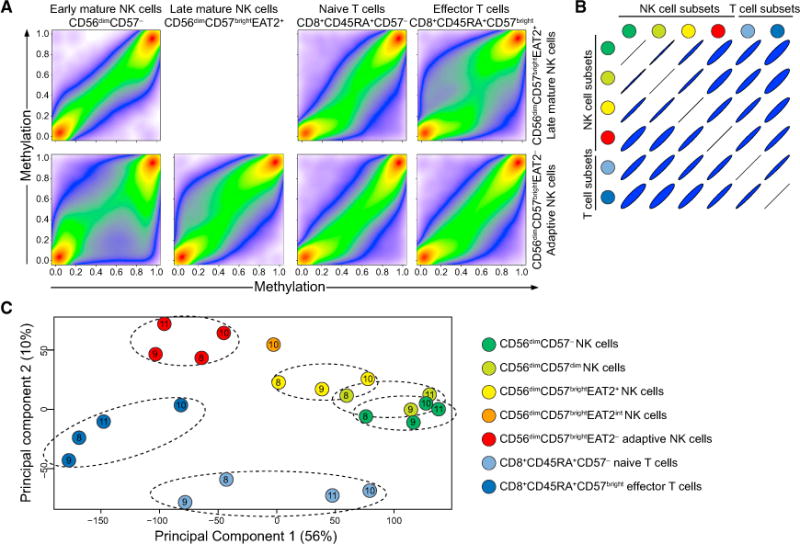
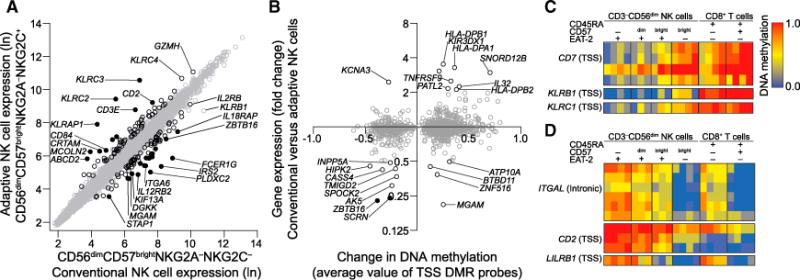
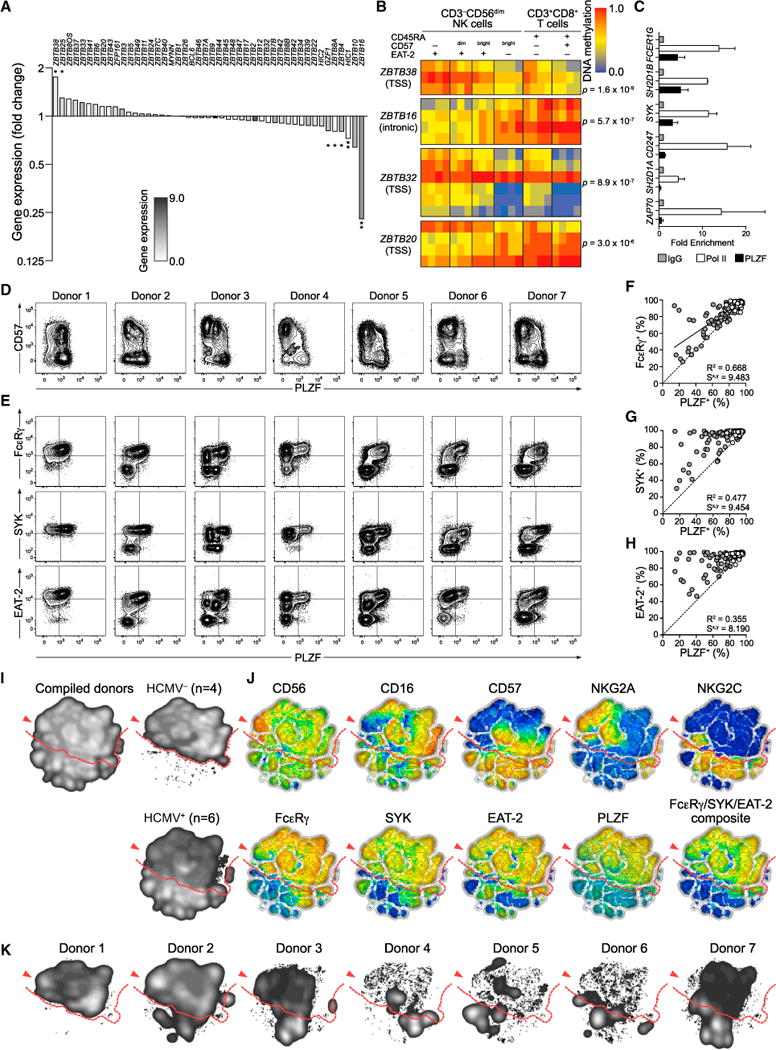
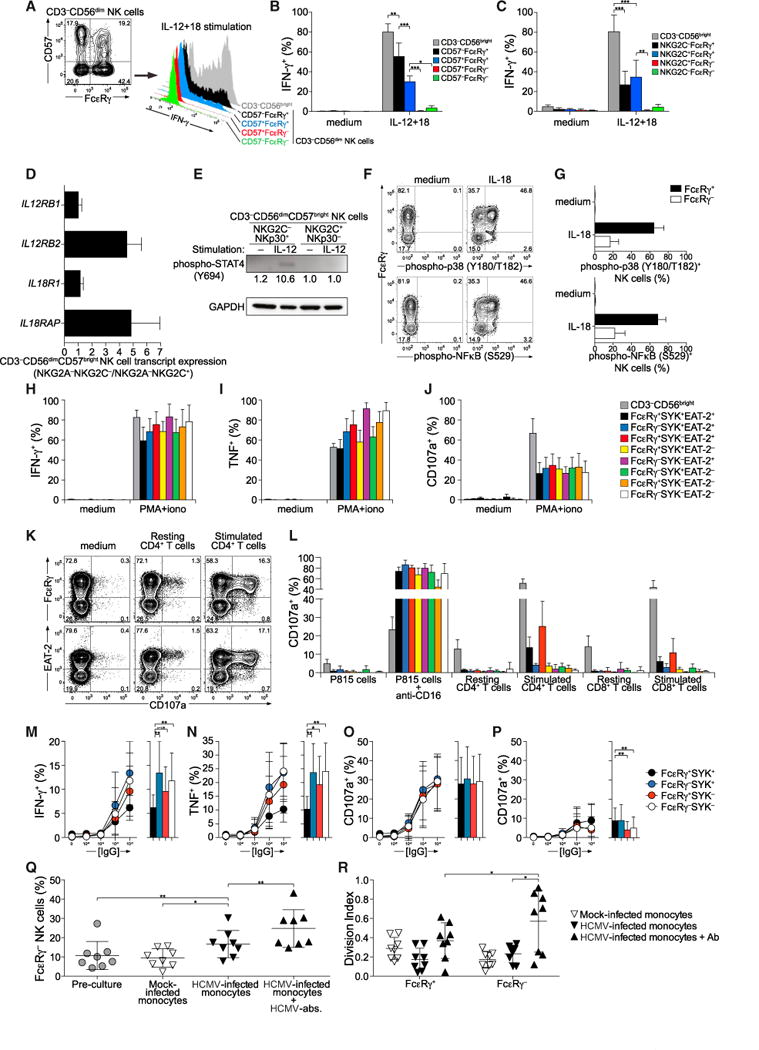
The mechanisms underlying human natural killer (NK) cell phenotypic and functional heterogeneity are unknown. Here, we describe the emergence of diverse subsets of human NK cells selectively lacking expression of signaling proteins after human cytomegalovirus (HCMV) infection. The absence of B and myeloid cell-related signaling protein expression in these NK cell subsets correlated with promoter DNA hypermethylation. Genome-wide DNA methylation patterns were strikingly similar between HCMV-associated adaptive NK cells and cytotoxic effector T cells but differed from those of canonical NK cells. Functional interrogation demonstrated altered cytokine responsiveness in adaptive NK cells that was linked to reduced expression of the transcription factor PLZF. Furthermore, subsets of adaptive NK cells demonstrated significantly reduced functional responses to activated autologous T cells. The present results uncover a spectrum of epigenetically unique adaptive NK cell subsets that diversify in response to viral infection and have distinct functional capabilities compared to canonical NK cell subsets.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures

Similar articles
-
Epigenetic modification and antibody-dependent expansion of memory-like NK cells in human cytomegalovirus-infected individuals.Immunity. 2015 Mar 17;42(3):431-42. doi: 10.1016/j.immuni.2015.02.013. Immunity. 2015. PMID: 25786175 Free PMC article.
-
Relationship of NKG2C Copy Number with the Distribution of Distinct Cytomegalovirus-Induced Adaptive NK Cell Subsets.J Immunol. 2016 May 1;196(9):3818-27. doi: 10.4049/jimmunol.1502438. Epub 2016 Mar 18. J Immunol. 2016. PMID: 26994220
-
Cytomegalovirus-Driven Adaption of Natural Killer Cells in NKG2Cnull Human Immunodeficiency Virus-Infected Individuals.Viruses. 2019 Mar 9;11(3):239. doi: 10.3390/v11030239. Viruses. 2019. PMID: 30857329 Free PMC article.
-
Adaptive reconfiguration of the human NK-cell compartment in response to cytomegalovirus: a different perspective of the host-pathogen interaction.Eur J Immunol. 2013 May;43(5):1133-41. doi: 10.1002/eji.201243117. Eur J Immunol. 2013. PMID: 23552990 Review.
-
Molecular characterization of HCMV-specific immune responses: Parallels between CD8(+) T cells, CD4(+) T cells, and NK cells.Eur J Immunol. 2015 Sep;45(9):2433-45. doi: 10.1002/eji.201545495. Epub 2015 Aug 24. Eur J Immunol. 2015. PMID: 26228786 Review.
Cited by
-
The transcription factor Bcl11b promotes both canonical and adaptive NK cell differentiation.Sci Immunol. 2021 Mar 12;6(57):eabc9801. doi: 10.1126/sciimmunol.abc9801. Sci Immunol. 2021. PMID: 33712472 Free PMC article.
-
Variations in killer-cell immunoglobulin-like receptor and human leukocyte antigen genes and immunity to malaria.Cell Mol Immunol. 2020 Aug;17(8):799-806. doi: 10.1038/s41423-020-0482-z. Epub 2020 Jun 15. Cell Mol Immunol. 2020. PMID: 32541835 Free PMC article. Review.
-
Harnessing CD16-Mediated NK Cell Functions to Enhance Therapeutic Efficacy of Tumor-Targeting mAbs.Cancers (Basel). 2021 May 20;13(10):2500. doi: 10.3390/cancers13102500. Cancers (Basel). 2021. PMID: 34065399 Free PMC article. Review.
-
Allogeneic natural killer cell therapy.Blood. 2023 Feb 23;141(8):856-868. doi: 10.1182/blood.2022016200. Blood. 2023. PMID: 36416736 Free PMC article. Review.
-
The novel mechanism facilitating chronic hepatitis B infection: immunometabolism and epigenetic modification reprogramming.Front Immunol. 2024 Jan 15;15:1349867. doi: 10.3389/fimmu.2024.1349867. eCollection 2024. Front Immunol. 2024. PMID: 38288308 Free PMC article. Review.
References
-
- Anfossi N, André P, Guia S, Falk CS, Roetynck S, Stewart CA, Breso V, Frassati C, Reviron D, Middleton D, et al. Human NK cell education by inhibitory receptors for MHC class I. Immunity. 2006;25:331–342. - PubMed
-
- Béziat V, Liu LL, Malmberg JA, Ivarsson MA, Sohlberg E, Björklund AT, Retière C, Sverremark-Ekström E, Traherne J, Ljungman P, et al. NK cell responses to cytomegalovirus infection lead to stable imprints in the human KIR repertoire and involve activating KIRs. Blood. 2013;121:2678–2688. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
