

K+ efflux agonists induce NLRP3 inflammasome activation independently of Ca2+ signaling
- PMID: 25762778
- PMCID: PMC4390495
- DOI: 10.4049/jimmunol.1402658
K+ efflux agonists induce NLRP3 inflammasome activation independently of Ca2+ signaling
Abstract
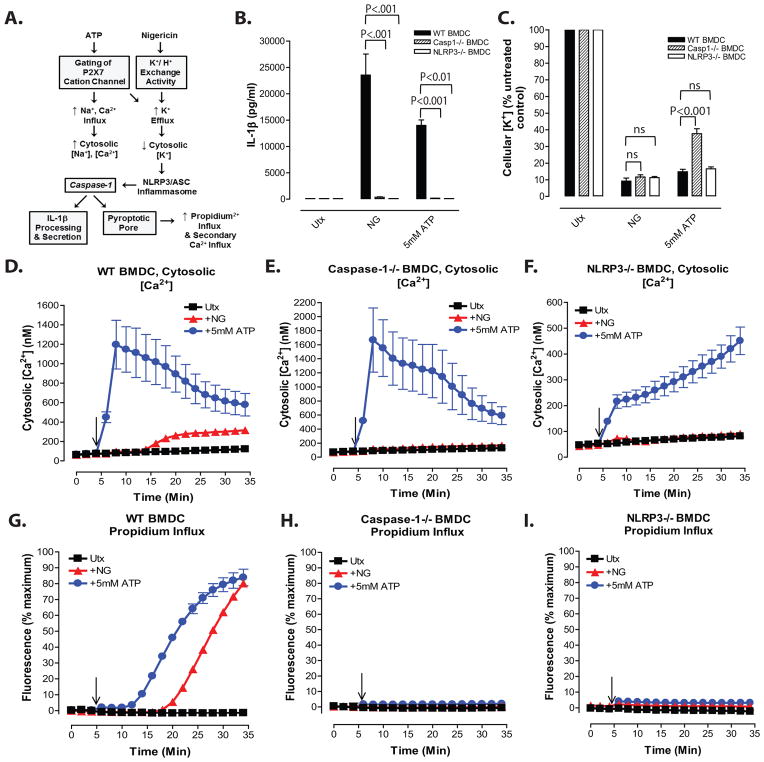
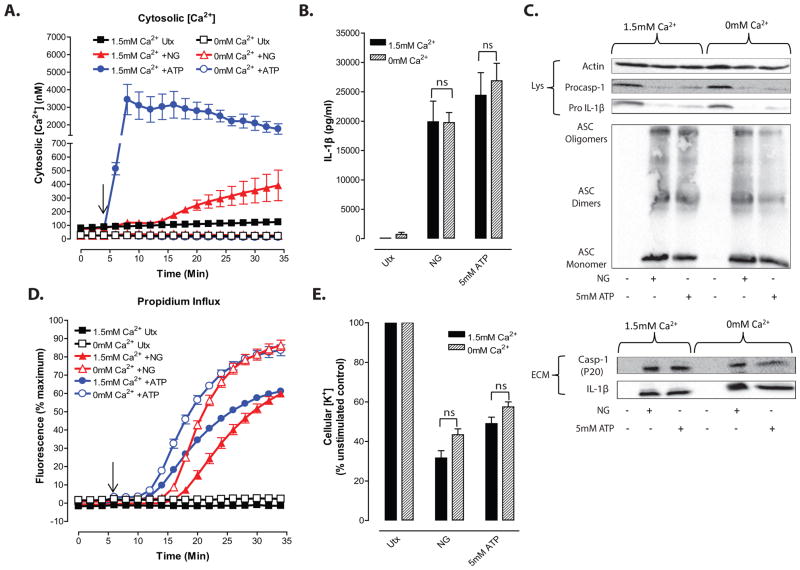
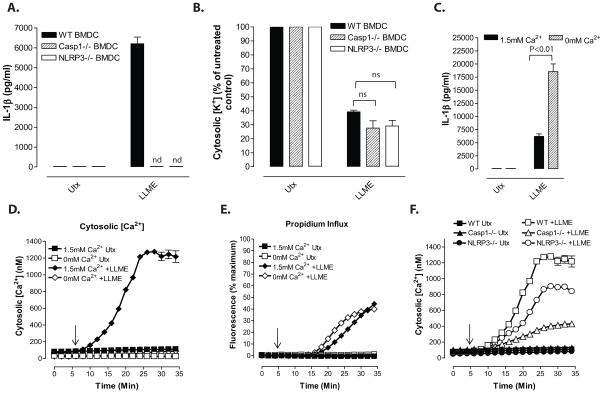
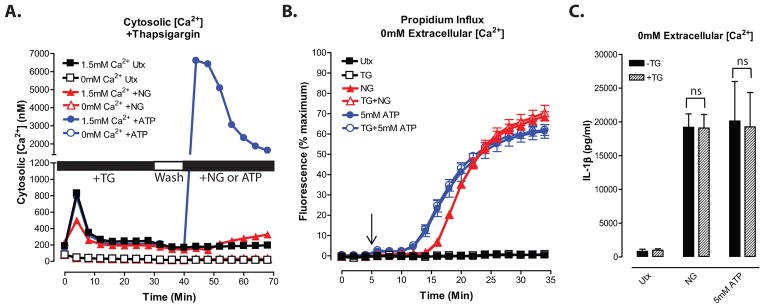
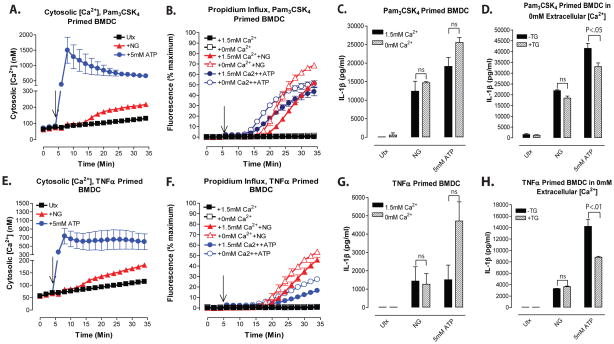
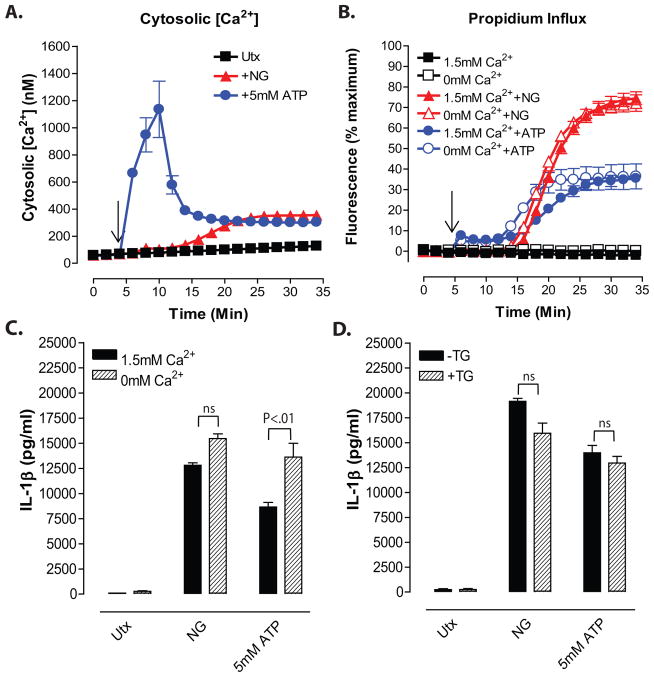
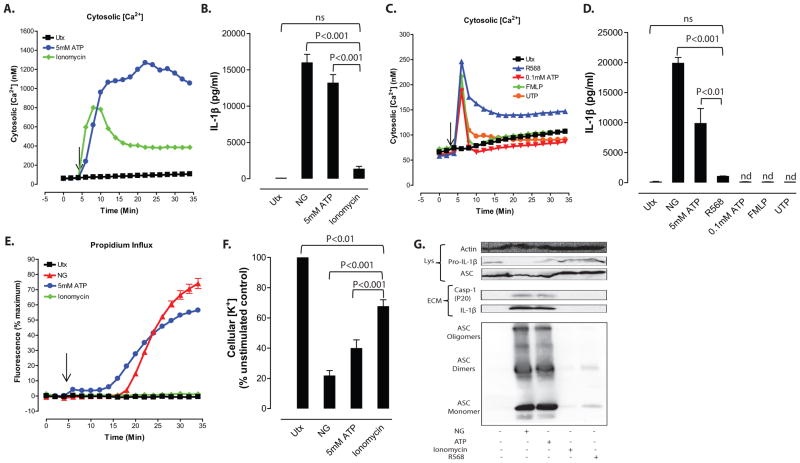
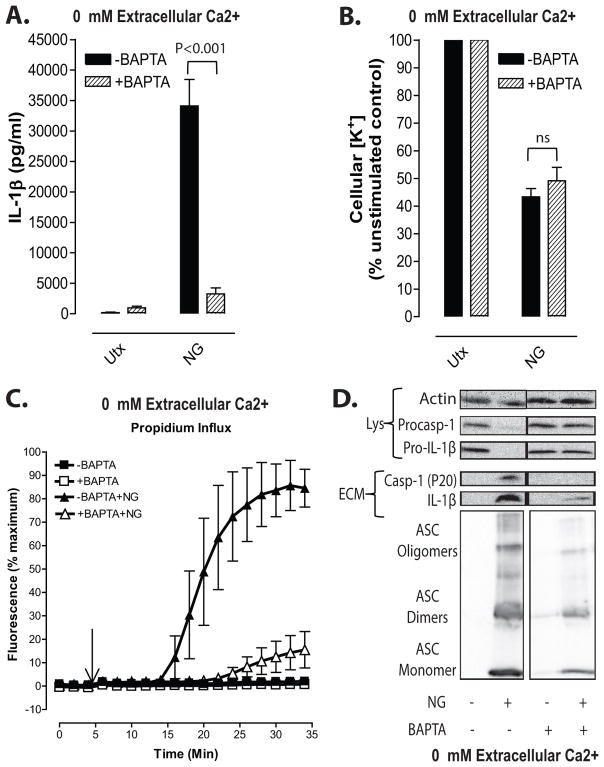
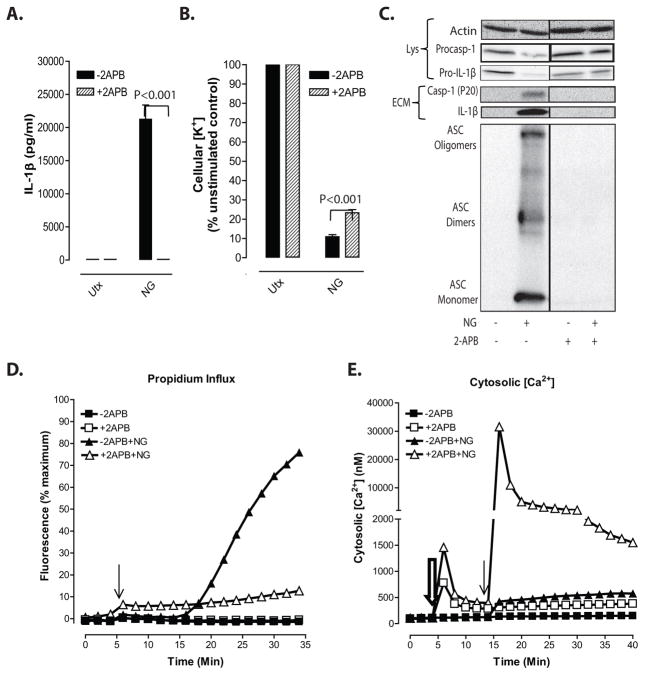
Perturbation of intracellular ion homeostasis is a major cellular stress signal for activation of NLRP3 inflammasome signaling that results in caspase-1-mediated production of IL-1β and pyroptosis. However, the relative contributions of decreased cytosolic K(+) concentration versus increased cytosolic Ca(2+) concentration ([Ca(2+)]) remain disputed and incompletely defined. We investigated roles for elevated cytosolic [Ca(2+)] in NLRP3 activation and downstream inflammasome signaling responses in primary murine dendritic cells and macrophages in response to two canonical NLRP3 agonists (ATP and nigericin) that facilitate primary K(+) efflux by mechanistically distinct pathways or the lysosome-destabilizing agonist Leu-Leu-O-methyl ester. The study provides three major findings relevant to this unresolved area of NLRP3 regulation. First, increased cytosolic [Ca(2+)] was neither a necessary nor sufficient signal for the NLRP3 inflammasome cascade during activation by endogenous ATP-gated P2X7 receptor channels, the exogenous bacterial ionophore nigericin, or the lysosomotropic agent Leu-Leu-O-methyl ester. Second, agonists for three Ca(2+)-mobilizing G protein-coupled receptors (formyl peptide receptor, P2Y2 purinergic receptor, and calcium-sensing receptor) expressed in murine dendritic cells were ineffective as activators of rapidly induced NLRP3 signaling when directly compared with the K(+) efflux agonists. Third, the intracellular Ca(2+) buffer, BAPTA, and the channel blocker, 2-aminoethoxydiphenyl borate, widely used reagents for disruption of Ca(2+)-dependent signaling pathways, strongly suppressed nigericin-induced NLRP3 inflammasome signaling via mechanisms dissociated from their canonical or expected effects on Ca(2+) homeostasis. The results indicate that the ability of K(+) efflux agonists to activate NLRP3 inflammasome signaling can be dissociated from changes in cytosolic [Ca(2+)] as a necessary or sufficient signal.
Copyright © 2015 by The American Association of Immunologists, Inc.
Figures
Similar articles
-
NLRP3 inflammasome signaling is activated by low-level lysosome disruption but inhibited by extensive lysosome disruption: roles for K+ efflux and Ca2+ influx.Am J Physiol Cell Physiol. 2016 Jul 1;311(1):C83-C100. doi: 10.1152/ajpcell.00298.2015. Epub 2016 May 11. Am J Physiol Cell Physiol. 2016. PMID: 27170638 Free PMC article.
-
Tripartite-motif protein 30 negatively regulates NLRP3 inflammasome activation by modulating reactive oxygen species production.J Immunol. 2010 Dec 15;185(12):7699-705. doi: 10.4049/jimmunol.1001099. Epub 2010 Nov 3. J Immunol. 2010. PMID: 21048113
-
Caspase-11 activates a canonical NLRP3 inflammasome by promoting K(+) efflux.Eur J Immunol. 2015 Oct;45(10):2927-36. doi: 10.1002/eji.201545772. Epub 2015 Aug 6. Eur J Immunol. 2015. PMID: 26173909
-
Potassium efflux fires the canon: Potassium efflux as a common trigger for canonical and noncanonical NLRP3 pathways.Eur J Immunol. 2015 Oct;45(10):2758-61. doi: 10.1002/eji.201545958. Epub 2015 Sep 21. Eur J Immunol. 2015. PMID: 26332156 Review.
-
Silica-induced inflammasome activation in macrophages: role of ATP and P2X7 receptor.Immunobiology. 2015 Sep;220(9):1101-6. doi: 10.1016/j.imbio.2015.05.004. Epub 2015 May 18. Immunobiology. 2015. PMID: 26024943 Review.
Cited by
-
Caspase-8 as an Effector and Regulator of NLRP3 Inflammasome Signaling.J Biol Chem. 2015 Aug 14;290(33):20167-84. doi: 10.1074/jbc.M115.652321. Epub 2015 Jun 22. J Biol Chem. 2015. PMID: 26100631 Free PMC article.
-
Unraveling the Role of the NLRP3 Inflammasome in Lymphoma: Implications in Pathogenesis and Therapeutic Strategies.Int J Mol Sci. 2024 Feb 17;25(4):2369. doi: 10.3390/ijms25042369. Int J Mol Sci. 2024. PMID: 38397043 Free PMC article. Review.
-
Apolipoprotein C3 induces inflammation and organ damage by alternative inflammasome activation.Nat Immunol. 2020 Jan;21(1):30-41. doi: 10.1038/s41590-019-0548-1. Epub 2019 Dec 9. Nat Immunol. 2020. PMID: 31819254
-
Anti-Inflammatory Therapeutic Mechanisms of Natural Products: Insight from Rosemary Diterpenes, Carnosic Acid and Carnosol.Biomedicines. 2023 Feb 13;11(2):545. doi: 10.3390/biomedicines11020545. Biomedicines. 2023. PMID: 36831081 Free PMC article. Review.
-
The pore-forming subunit Kir6.1 of the K-ATP channel negatively regulates the NLRP3 inflammasome to control insulin resistance by interacting with NLRP3.Exp Mol Med. 2019 Aug 6;51(8):1-13. doi: 10.1038/s12276-019-0291-6. Exp Mol Med. 2019. PMID: 31387986 Free PMC article.
References
-
- Warren J. Williams Hematology. 8. McGraw Hill Companies; 2010. The Inflammatory Response. Access Medicine. Web. 7 August 2014.
-
- Schroder K, Tschopp J. The Inflammasomes. Cell. 2010;140(6):821–832. - PubMed
-
- Gross O, Thomas CJ, Guarda G, Tschopp J. The inflammasome: an integrated view. Immunological Reviews. 2011;243(1):136–151. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous