

Angiopoietin-1 requires oxidant signaling through p47phox to promote endothelial barrier defense
- PMID: 25761062
- PMCID: PMC4356555
- DOI: 10.1371/journal.pone.0119577
Angiopoietin-1 requires oxidant signaling through p47phox to promote endothelial barrier defense
Abstract
Background: Reactive oxygen species (ROS) are largely considered to be pathogenic to normal endothelial function in disease states such as sepsis. We hypothesized that Angiopoietin-1 (Angpt-1), an endogenous agonist of the endothelial-specific receptor, Tie-2, promotes barrier defense by activating NADPH oxidase (NOX) signaling.
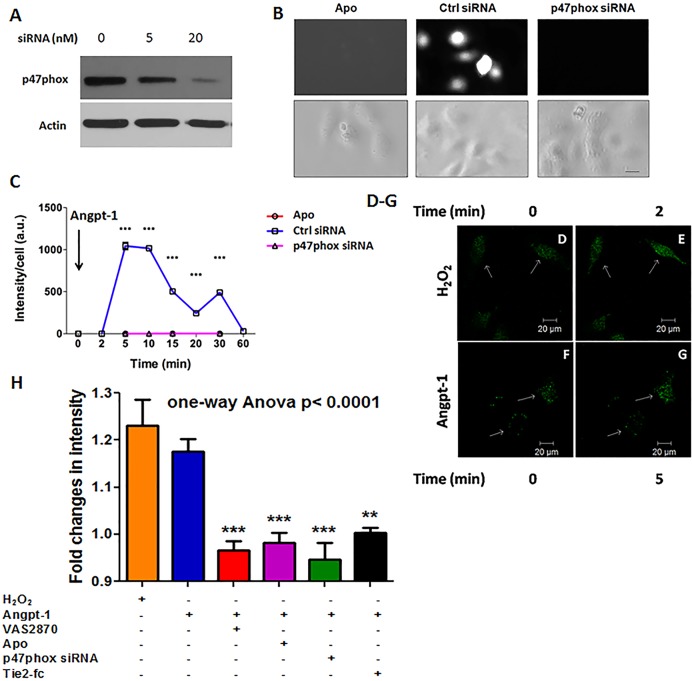
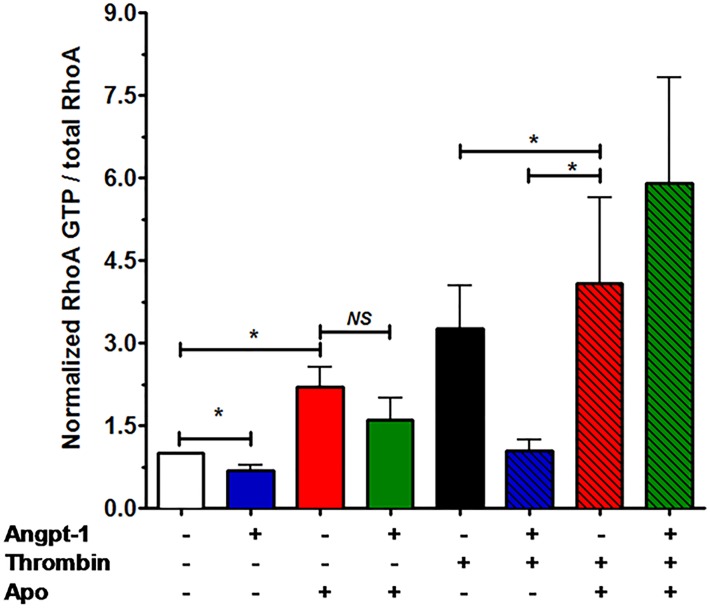
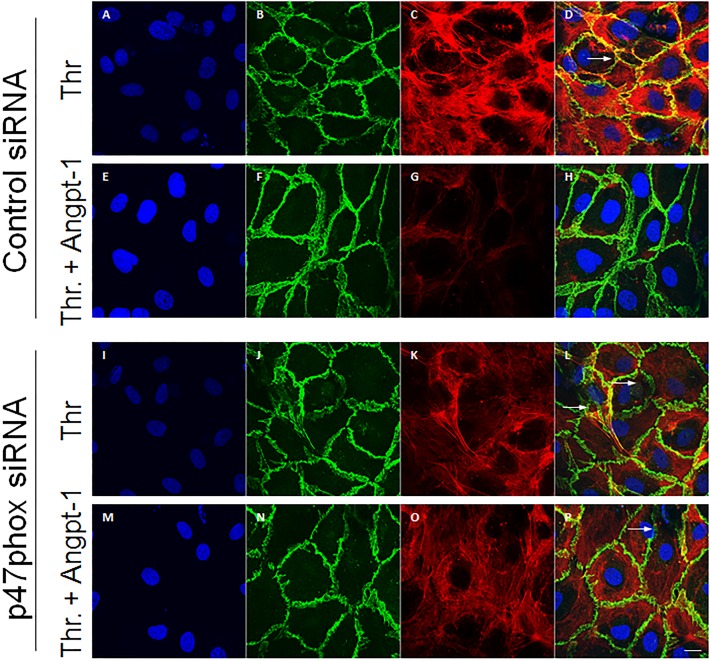
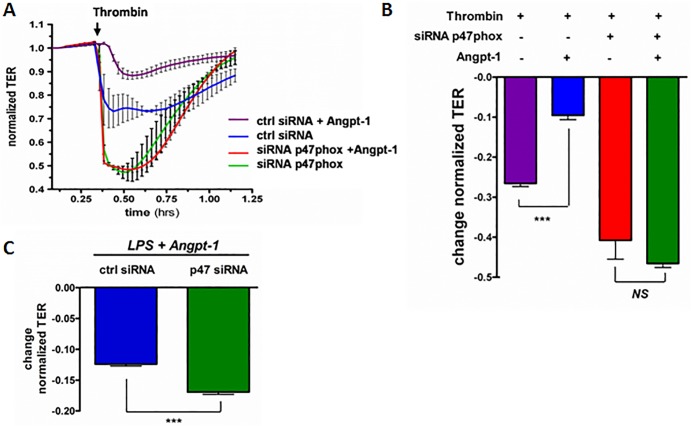
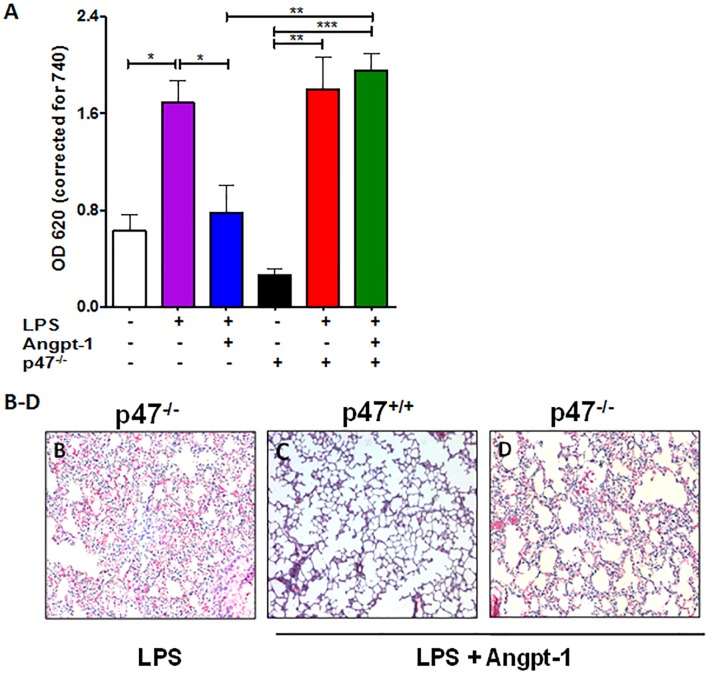
Methods and findings: Using primary human microvascular endothelial cells (HMVECs), we found that Angpt-1 stimulation induces phosphorylation of p47phox and a brief oxidative burst that is lost when chemical inhibitors of NOX activity or siRNA against the NOX component p47phox were applied. As a result, there was attenuated ROS activity, disrupted junctional contacts, enhanced actin stress fiber accumulation, and induced gap formation between confluent HMVECs. All of these changes were associated with weakened barrier function. The ability of Angpt-1 to prevent identical changes induced by inflammatory permeability mediators, thrombin and lipopolysaccharides (LPS), was abrogated by p47phox knockdown. P47phox was required for Angpt-1 to activate Rac1 and inhibit mediator-induced activation of the small GTPase RhoA. Finally, Angpt-1 gene transfer prevented vascular leakage in wildtype mice exposed to systemically administered LPS, but not in p47phox knock out (p47-/-) littermates.
Conclusions: These results suggest an essential role for NOX signaling in Angpt-1-mediated endothelial barrier defense against mediators of systemic inflammation. More broadly, oxidants generated for signal transduction may have a barrier-promoting role in vascular endothelium.
Conflict of interest statement
Figures
Similar articles
-
Angiopoietin-1 requires IQ domain GTPase-activating protein 1 to activate Rac1 and promote endothelial barrier defense.Arterioscler Thromb Vasc Biol. 2011 Nov;31(11):2643-52. doi: 10.1161/ATVBAHA.111.233189. Arterioscler Thromb Vasc Biol. 2011. PMID: 21885850 Free PMC article.
-
Phosphoinositide 3-kinase mediates CD40 ligand-induced oxidative stress and endothelial dysfunction via Rac1 and NADPH oxidase 2.J Thromb Haemost. 2010 Feb;8(2):397-406. doi: 10.1111/j.1538-7836.2009.03683.x. Epub 2009 Nov 6. J Thromb Haemost. 2010. PMID: 19895673
-
Blockade of NOX2 and STIM1 signaling limits lipopolysaccharide-induced vascular inflammation.J Clin Invest. 2013 Feb;123(2):887-902. doi: 10.1172/JCI65647. Epub 2013 Jan 25. J Clin Invest. 2013. PMID: 23348743 Free PMC article.
-
Reactive oxygen species as mediators of angiogenesis signaling: role of NAD(P)H oxidase.Mol Cell Biochem. 2004 Sep;264(1-2):85-97. doi: 10.1023/b:mcbi.0000044378.09409.b5. Mol Cell Biochem. 2004. PMID: 15544038 Review.
-
Organizers and activators: Cytosolic Nox proteins impacting on vascular function.Free Radic Biol Med. 2017 Aug;109:22-32. doi: 10.1016/j.freeradbiomed.2017.03.017. Epub 2017 Mar 21. Free Radic Biol Med. 2017. PMID: 28336130 Review.
Cited by
-
Idiopathic systemic capillary leak syndrome (Clarkson disease).J Allergy Clin Immunol. 2017 Sep;140(3):663-670. doi: 10.1016/j.jaci.2016.10.042. Epub 2016 Dec 22. J Allergy Clin Immunol. 2017. PMID: 28012935 Free PMC article. Review.
-
Flavone inhibits migration through DLC1/RhoA pathway by decreasing ROS generation in breast cancer cells.In Vitro Cell Dev Biol Anim. 2016 May;52(5):589-97. doi: 10.1007/s11626-016-0010-8. Epub 2016 Mar 2. In Vitro Cell Dev Biol Anim. 2016. PMID: 26935193
-
Calpain-14 and its association with eosinophilic esophagitis.J Allergy Clin Immunol. 2017 Jun;139(6):1762-1771.e7. doi: 10.1016/j.jaci.2016.09.027. Epub 2017 Jan 25. J Allergy Clin Immunol. 2017. PMID: 28131390 Free PMC article. Review.
-
Multiplexed, high-throughput measurements of cell contraction and endothelial barrier function.Lab Invest. 2019 Jan;99(1):138-145. doi: 10.1038/s41374-018-0136-2. Epub 2018 Oct 11. Lab Invest. 2019. PMID: 30310180 Free PMC article.
-
The Angiopoietin-Tie2 Pathway in Critical Illness.Crit Care Clin. 2020 Apr;36(2):201-216. doi: 10.1016/j.ccc.2019.12.003. Epub 2020 Jan 31. Crit Care Clin. 2020. PMID: 32172809 Free PMC article. Review.
References
-
- Dumont DJ, Gradwohl G, Fong GH, Puri MC, Gertsenstein M, et al. (1994) Dominant-negative and targeted null mutations in the endothelial receptor tyrosine kinase, tek, reveal a critical role in vasculogenesis of the embryo. Genes Dev 8: 1897–1909. - PubMed
-
- Suri C, Jones PF, Patan S, Bartunkova S, Maisonpierre PC, et al. (1996) Requisite role of angiopoietin-1, a ligand for the TIE2 receptor, during embryonic angiogenesis. Cell 87: 1171–1180. - PubMed
-
- Wong AL, Haroon ZA, Werner S, Dewhirst MW, Greenberg CS, et al. (1997) Tie2 expression and phosphorylation in angiogenic and quiescent adult tissues. Circ Res 81: 567–574. - PubMed
-
- Thurston G, Rudge JS, Ioffe E, Zhou H, Ross L, et al. (2000) Angiopoietin-1 protects the adult vasculature against plasma leakage. Nat Med 6: 460–463. - PubMed
-
- Thurston G, Suri C, Smith K, McClain J, Sato TN, et al. (1999) Leakage-resistant blood vessels in mice transgenically overexpressing angiopoietin-1. Science 286: 2511–2514. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
