

Lin28b promotes fetal B lymphopoiesis through the transcription factor Arid3a
- PMID: 25753579
- PMCID: PMC4387290
- DOI: 10.1084/jem.20141510
Lin28b promotes fetal B lymphopoiesis through the transcription factor Arid3a
Abstract
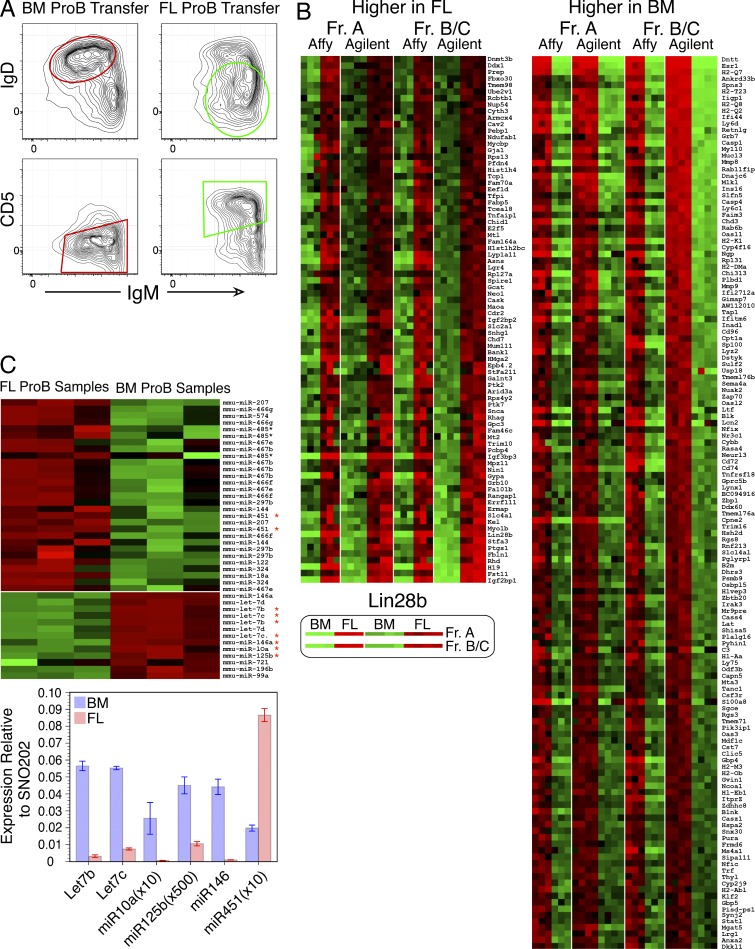
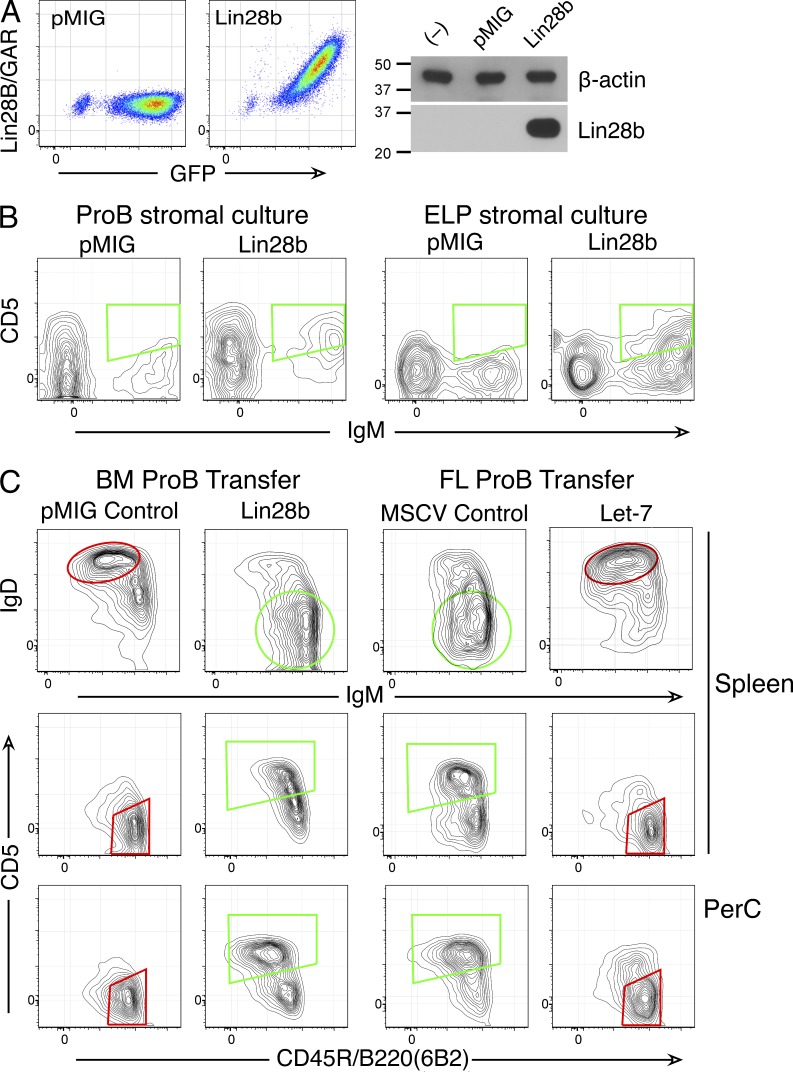
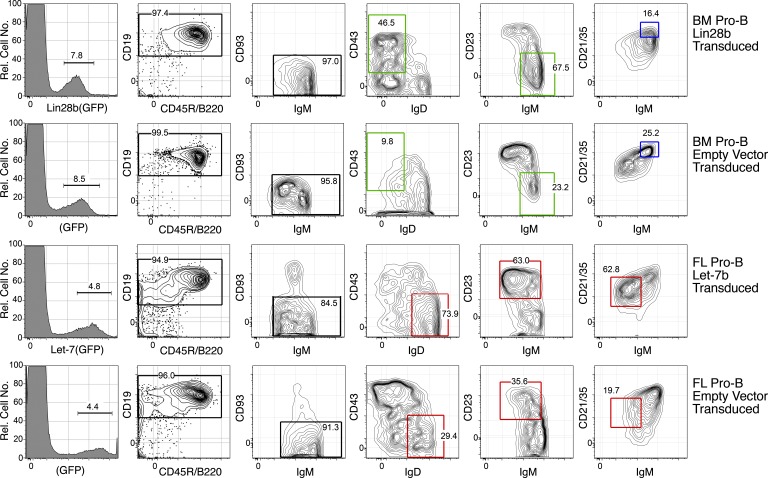
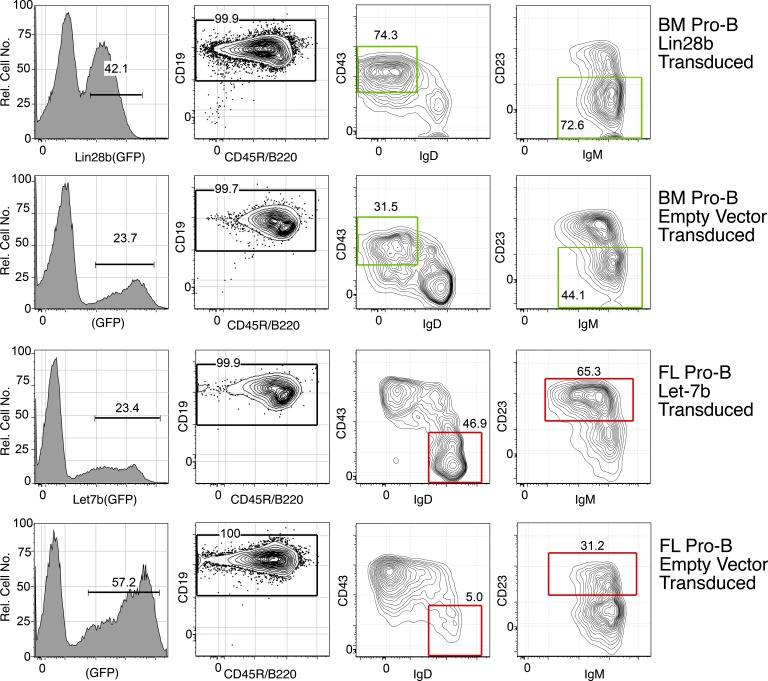
Mouse B cell precursors from fetal liver and adult bone marrow (BM) generate distinctive B cell progeny when transplanted into immunodeficient recipients, supporting a two-pathway model for B lymphopoiesis, fetal "B-1" and adult "B-2." Recently, Lin28b was shown to be important for the switch between fetal and adult pathways; however, neither the mechanism of Lin28b action nor the importance of B cell antigen receptor (BCR) signaling in this process was addressed. Here, we report key advances in our understanding of the regulation of B-1/B-2 development. First, modulation of Let-7 in fetal pro-B cells is sufficient to alter fetal B-1 development to produce B cells resembling the progeny of adult B-2 development. Second, intact BCR signaling is required for the generation of B1a B cells from Lin28b-transduced BM progenitors, supporting a requirement for ligand-dependent selection, as is the case for normal B1a B cells. Third, the VH repertoire of Lin28b-induced BM B1a B cells differs from that of normal B1a, suggesting persisting differences from fetal progenitors. Finally, we identify the Arid3a transcription factor as a key target of Let-7, whose ectopic expression is sufficient to induce B-1 development in adult pro-B cells and whose silencing by knockdown blocks B-1 development in fetal pro-B cells.
© 2015 Zhou et al.
Figures
Similar articles
-
Crucial Role of Increased Arid3a at the Pre-B and Immature B Cell Stages for B1a Cell Generation.Front Immunol. 2019 Mar 15;10:457. doi: 10.3389/fimmu.2019.00457. eCollection 2019. Front Immunol. 2019. PMID: 30930899 Free PMC article.
-
Perspectives on fetal derived CD5+ B1 B cells.Eur J Immunol. 2015 Nov;45(11):2978-84. doi: 10.1002/eji.201445146. Epub 2015 Sep 23. Eur J Immunol. 2015. PMID: 26339791 Free PMC article. Review.
-
A developmental switch between fetal and adult B lymphopoiesis.Ann N Y Acad Sci. 2015 Dec;1362:8-15. doi: 10.1111/nyas.12769. Epub 2015 Apr 30. Ann N Y Acad Sci. 2015. PMID: 25931205 Review.
-
Distinct Genetic Networks Orchestrate the Emergence of Specific Waves of Fetal and Adult B-1 and B-2 Development.Immunity. 2016 Sep 20;45(3):527-539. doi: 10.1016/j.immuni.2016.07.012. Epub 2016 Aug 23. Immunity. 2016. PMID: 27566938 Free PMC article.
-
Lin28b reprograms adult bone marrow hematopoietic progenitors to mediate fetal-like lymphopoiesis.Science. 2012 Mar 9;335(6073):1195-200. doi: 10.1126/science.1216557. Epub 2012 Feb 16. Science. 2012. PMID: 22345399 Free PMC article.
Cited by
-
B-1 Cell Heterogeneity and the Regulation of Natural and Antigen-Induced IgM Production.Front Immunol. 2016 Sep 9;7:324. doi: 10.3389/fimmu.2016.00324. eCollection 2016. Front Immunol. 2016. PMID: 27667991 Free PMC article. Review.
-
Cell Cycle Regulation of Stem Cells by MicroRNAs.Stem Cell Rev Rep. 2018 Jun;14(3):309-322. doi: 10.1007/s12015-018-9808-y. Stem Cell Rev Rep. 2018. PMID: 29541978 Free PMC article. Review.
-
Ezh2 Represses Transcription of Innate Lymphoid Genes in B Lymphocyte Progenitors and Maintains the B-2 Cell Fate.J Immunol. 2020 Apr 1;204(7):1760-1769. doi: 10.4049/jimmunol.1901188. Epub 2020 Feb 24. J Immunol. 2020. PMID: 32094206 Free PMC article.
-
MicroRNAs in Control of Stem Cells in Normal and Malignant Hematopoiesis.Curr Stem Cell Rep. 2016 Sep;2(3):183-196. doi: 10.1007/s40778-016-0057-1. Epub 2016 Jul 1. Curr Stem Cell Rep. 2016. PMID: 27547713 Free PMC article.
-
MicroRNA-regulated B cells in obesity.Immunometabolism (Cobham). 2022 Aug 5;4(3):e00005. doi: 10.1097/IN9.0000000000000005. eCollection 2022 Jul. Immunometabolism (Cobham). 2022. PMID: 35966635 Free PMC article. Review.
References
-
- Goebel P., Montalbano A., Ayers N., Kompfner E., Dickinson L., Webb C.F., and Feeney A.J.. 2002. High frequency of matrix attachment regions and cut-like protein x/CCAAT-displacement protein and B cell regulator of IgH transcription binding sites flanking Ig V region genes. J. Immunol. 169:2477–2487 10.4049/jimmunol.169.5.2477 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
