

Sex hormones affect neurotransmitters and shape the adult female brain during hormonal transition periods
- PMID: 25750611
- PMCID: PMC4335177
- DOI: 10.3389/fnins.2015.00037
Sex hormones affect neurotransmitters and shape the adult female brain during hormonal transition periods
Abstract
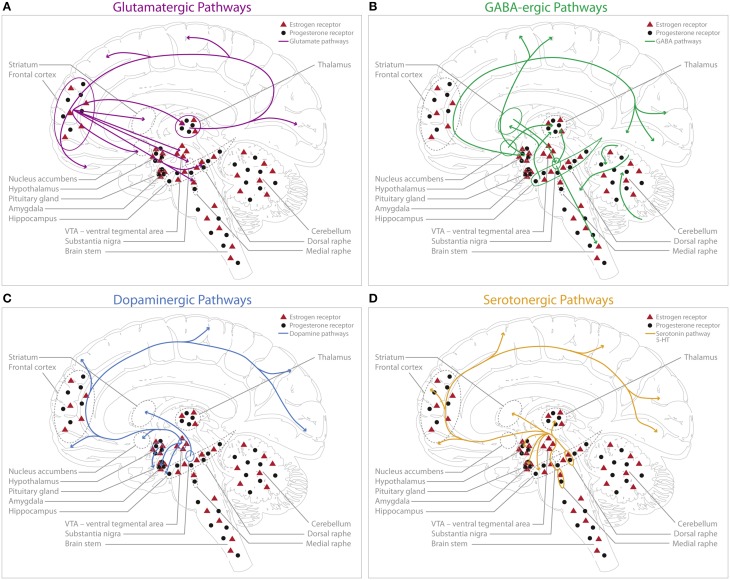
Sex hormones have been implicated in neurite outgrowth, synaptogenesis, dendritic branching, myelination and other important mechanisms of neural plasticity. Here we review the evidence from animal experiments and human studies reporting interactions between sex hormones and the dominant neurotransmitters, such as serotonin, dopamine, GABA and glutamate. We provide an overview of accumulating data during physiological and pathological conditions and discuss currently conceptualized theories on how sex hormones potentially trigger neuroplasticity changes through these four neurochemical systems. Many brain regions have been demonstrated to express high densities for estrogen- and progesterone receptors, such as the amygdala, the hypothalamus, and the hippocampus. As the hippocampus is of particular relevance in the context of mediating structural plasticity in the adult brain, we put particular emphasis on what evidence could be gathered thus far that links differences in behavior, neurochemical patterns and hippocampal structure to a changing hormonal environment. Finally, we discuss how physiologically occurring hormonal transition periods in humans can be used to model how changes in sex hormones influence functional connectivity, neurotransmission and brain structure in vivo.
Keywords: estrogens; hormonal transition periods; neurotransmitters; plasticity; progesterone.
Figures

Similar articles
-
Optimizing quality of life through sex steroids by their effects on neurotransmitters.Climacteric. 2019 Feb;22(1):55-59. doi: 10.1080/13697137.2018.1543265. Epub 2018 Dec 20. Climacteric. 2019. PMID: 30570355 Review.
-
Estrogen- and progesterone-mediated structural neuroplasticity in women: evidence from neuroimaging.Brain Struct Funct. 2016 Nov;221(8):3845-3867. doi: 10.1007/s00429-016-1197-x. Epub 2016 Feb 20. Brain Struct Funct. 2016. PMID: 26897178 Free PMC article. Review.
-
Enriched environment influences hormonal status and hippocampal brain derived neurotrophic factor in a sex dependent manner.Neuroscience. 2009 Dec 1;164(2):788-97. doi: 10.1016/j.neuroscience.2009.08.054. Epub 2009 Aug 29. Neuroscience. 2009. PMID: 19723563
-
Interactions between hormones and epilepsy.Seizure. 2015 May;28:3-11. doi: 10.1016/j.seizure.2015.02.012. Epub 2015 Feb 23. Seizure. 2015. PMID: 25765693 Review.
-
Steroid-induced hippocampal synaptic plasticity: sex differences and similarities.Neuron Glia Biol. 2004 Aug;1(3):219-29. doi: 10.1017/S1740925X05000086. Neuron Glia Biol. 2004. PMID: 18634599
Cited by
-
Hypothalamic-Pituitary-End-Organ Axes: Hormone Function in Female Patients with Major Depressive Disorder.Neurosci Bull. 2021 Aug;37(8):1176-1187. doi: 10.1007/s12264-021-00689-6. Epub 2021 Apr 28. Neurosci Bull. 2021. PMID: 33909242 Free PMC article. Review.
-
Gender differences in factors associated with symptoms of depression among high school students: an examination of the direct and indirect effects of insomnia symptoms and physical activity.Health Psychol Behav Med. 2019 May 15;7(1):179-192. doi: 10.1080/21642850.2019.1615926. Health Psychol Behav Med. 2019. PMID: 34040846 Free PMC article.
-
The attention-emotion interaction in healthy female participants on oral contraceptives during 1-week escitalopram intake.Front Neurosci. 2022 Sep 9;16:809269. doi: 10.3389/fnins.2022.809269. eCollection 2022. Front Neurosci. 2022. PMID: 36161146 Free PMC article.
-
Epigenetic mechanisms underlying sex differences in the brain and behavior.Trends Neurosci. 2024 Jan;47(1):18-35. doi: 10.1016/j.tins.2023.09.007. Epub 2023 Nov 15. Trends Neurosci. 2024. PMID: 37968206 Free PMC article. Review.
-
Menstrual cycle-driven hormone concentrations co-fluctuate with white and gray matter architecture changes across the whole brain.Hum Brain Mapp. 2024 Aug 1;45(11):e26785. doi: 10.1002/hbm.26785. Hum Brain Mapp. 2024. PMID: 39031470 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials