

Non-redundant and redundant roles of cytomegalovirus gH/gL complexes in host organ entry and intra-tissue spread
- PMID: 25659098
- PMCID: PMC4450070
- DOI: 10.1371/journal.ppat.1004640
Non-redundant and redundant roles of cytomegalovirus gH/gL complexes in host organ entry and intra-tissue spread
Abstract
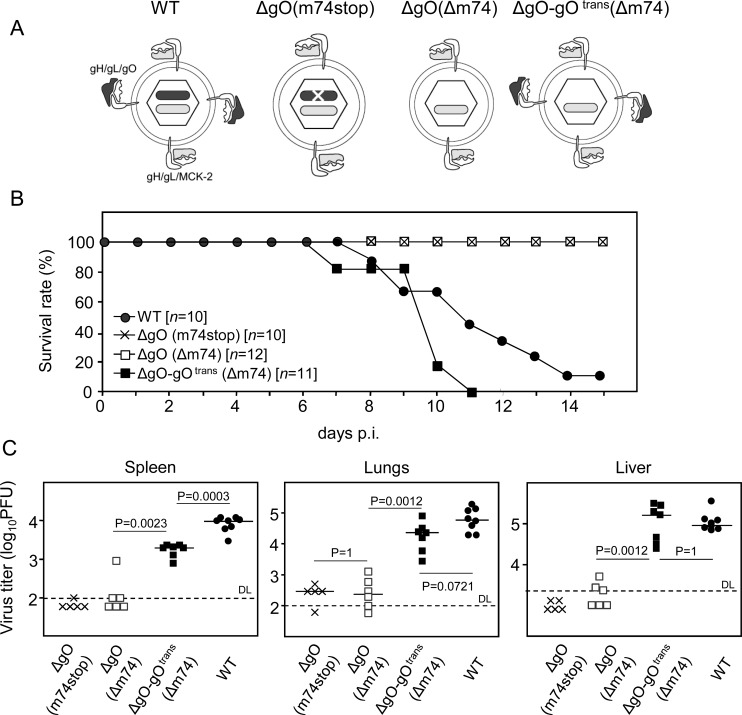
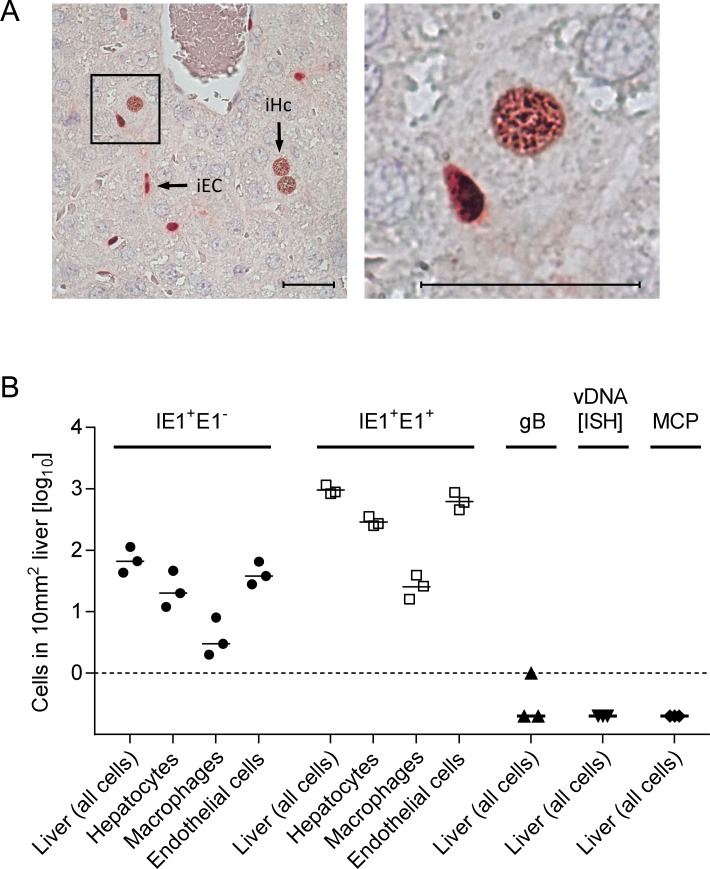
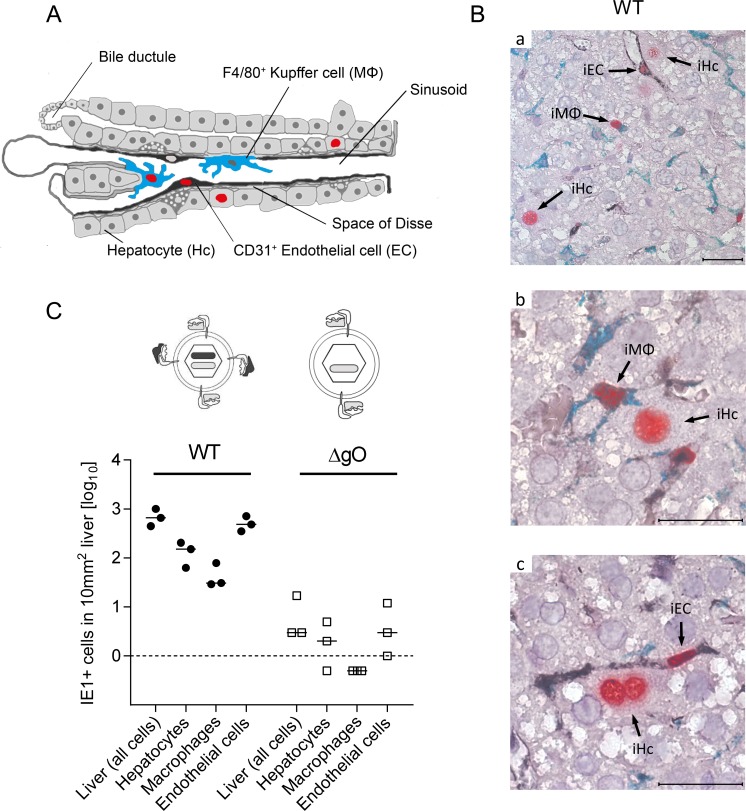
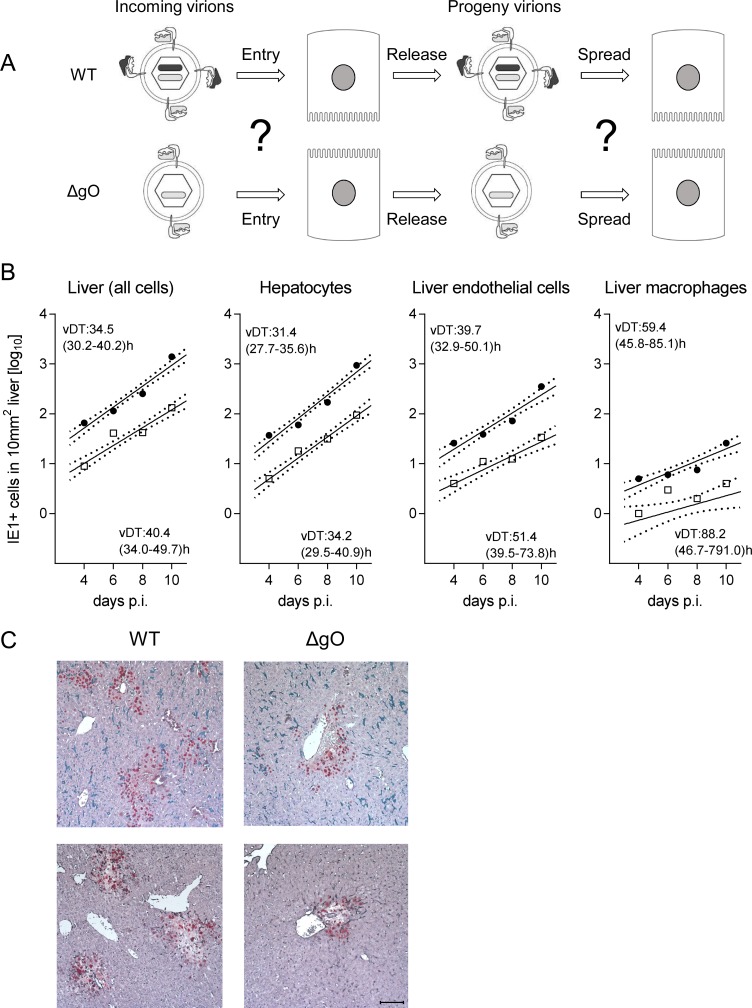
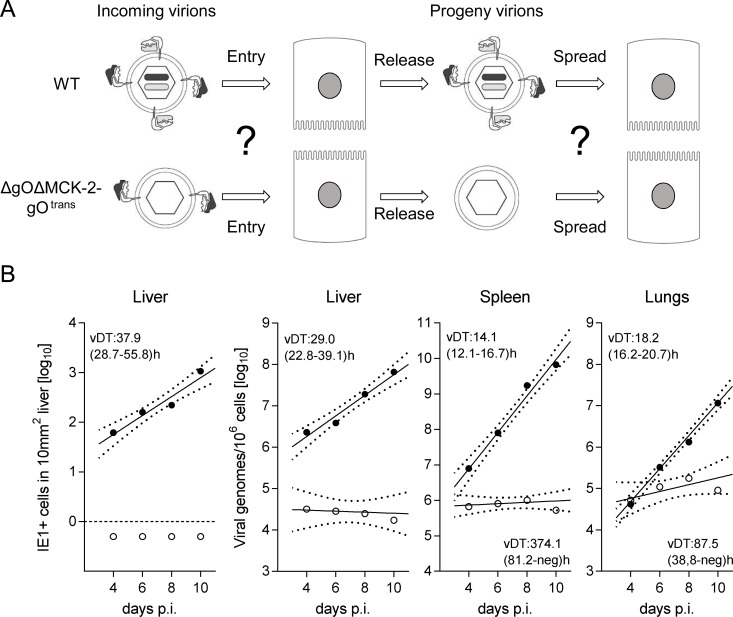
Herpesviruses form different gH/gL virion envelope glycoprotein complexes that serve as entry complexes for mediating viral cell-type tropism in vitro; their roles in vivo, however, remained speculative and can be addressed experimentally only in animal models. For murine cytomegalovirus two alternative gH/gL complexes, gH/gL/gO and gH/gL/MCK-2, have been identified. A limitation of studies on viral tropism in vivo has been the difficulty in distinguishing between infection initiation by viral entry into first-hit target cells and subsequent cell-to-cell spread within tissues. As a new strategy to dissect these two events, we used a gO-transcomplemented ΔgO mutant for providing the gH/gL/gO complex selectively for the initial entry step, while progeny virions lack gO in subsequent rounds of infection. Whereas gH/gL/gO proved to be critical for establishing infection by efficient entry into diverse cell types, including liver macrophages, endothelial cells, and hepatocytes, it was dispensable for intra-tissue spread. Notably, the salivary glands, the source of virus for host-to-host transmission, represent an exception in that entry into virus-producing cells did not strictly depend on either the gH/gL/gO or the gH/gL/MCK-2 complex. Only if both complexes were absent in gO and MCK-2 double-knockout virus, in vivo infection was abolished at all sites.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
Pathogen at the Gates: Human Cytomegalovirus Entry and Cell Tropism.Viruses. 2018 Dec 11;10(12):704. doi: 10.3390/v10120704. Viruses. 2018. PMID: 30544948 Free PMC article. Review.
-
Principles for studying in vivo attenuation of virus mutants: defining the role of the cytomegalovirus gH/gL/gO complex as a paradigm.Med Microbiol Immunol. 2015 Jun;204(3):295-305. doi: 10.1007/s00430-015-0405-2. Epub 2015 Mar 18. Med Microbiol Immunol. 2015. PMID: 25782576 Review.
-
Human Cytomegalovirus gH/gL/gO Promotes the Fusion Step of Entry into All Cell Types, whereas gH/gL/UL128-131 Broadens Virus Tropism through a Distinct Mechanism.J Virol. 2015 Sep;89(17):8999-9009. doi: 10.1128/JVI.01325-15. Epub 2015 Jun 17. J Virol. 2015. PMID: 26085146 Free PMC article.
-
A viral regulator of glycoprotein complexes contributes to human cytomegalovirus cell tropism.Proc Natl Acad Sci U S A. 2015 Apr 7;112(14):4471-6. doi: 10.1073/pnas.1419875112. Epub 2015 Mar 23. Proc Natl Acad Sci U S A. 2015. PMID: 25831500 Free PMC article.
-
The viral chemokine MCK-2 of murine cytomegalovirus promotes infection as part of a gH/gL/MCK-2 complex.PLoS Pathog. 2013;9(7):e1003493. doi: 10.1371/journal.ppat.1003493. Epub 2013 Jul 25. PLoS Pathog. 2013. PMID: 23935483 Free PMC article.
Cited by
-
Persistent viral replication and the development of T-cell responses after intranasal infection by MCMV.Med Microbiol Immunol. 2019 Aug;208(3-4):457-468. doi: 10.1007/s00430-019-00589-7. Epub 2019 Mar 8. Med Microbiol Immunol. 2019. PMID: 30848361 Free PMC article.
-
The pentameric complex drives immunologically covert cell-cell transmission of wild-type human cytomegalovirus.Proc Natl Acad Sci U S A. 2017 Jun 6;114(23):6104-6109. doi: 10.1073/pnas.1704809114. Epub 2017 May 22. Proc Natl Acad Sci U S A. 2017. PMID: 28533400 Free PMC article.
-
Cytomegalovirus-induced peroxynitrite promotes virus entry and contributes to pathogenesis in a murine model of infection.mBio. 2024 Aug 14;15(8):e0315223. doi: 10.1128/mbio.03152-23. Epub 2024 Jul 2. mBio. 2024. PMID: 38953361 Free PMC article.
-
Pathogen at the Gates: Human Cytomegalovirus Entry and Cell Tropism.Viruses. 2018 Dec 11;10(12):704. doi: 10.3390/v10120704. Viruses. 2018. PMID: 30544948 Free PMC article. Review.
-
Editorship for Medical Microbiology and Immunology: the baton has been passed on.Med Microbiol Immunol. 2018 Feb;207(1):1-2. doi: 10.1007/s00430-017-0533-y. Med Microbiol Immunol. 2018. PMID: 29313096 No abstract available.
References
-
- Pedersen SM Hollsberg P (2006) Complexities in human herpesvirus-6A and-6B binding to host cells. Virology 356:1–3. - PubMed
-
- Adler B, Sinzger C (2013) Cytomegalovirus inter-strain variance in cell-type tropism. In: Reddehase, editor. Cytomegaloviruses: from molecular pathogenesis to intervention. Caister Academic Press, Norfolk, UK: Vol. II, pp. 297–322.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
