

doi: 10.1128/JVI.02677-14.
Epub 2015 Jan 14.
Overlapping local and long-range RNA-RNA interactions modulate dengue virus genome cyclization and replication
Affiliations
- PMID: 25589642
- PMCID: PMC4337542
- DOI: 10.1128/JVI.02677-14
Item in Clipboard
Overlapping local and long-range RNA-RNA interactions modulate dengue virus genome cyclization and replication
J Virol.
2015 Mar.
Abstract
The dengue virus genome is a dynamic molecule that adopts different conformations in the infected cell. Here, using RNA folding predictions, chemical probing analysis, RNA binding assays, and functional studies, we identified new cis-acting elements present in the capsid coding sequence that facilitate cyclization of the viral RNA by hybridization with a sequence involved in a local dumbbell structure at the viral 3' untranslated region (UTR). The identified interaction differentially enhances viral replication in mosquito and mammalian cells.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
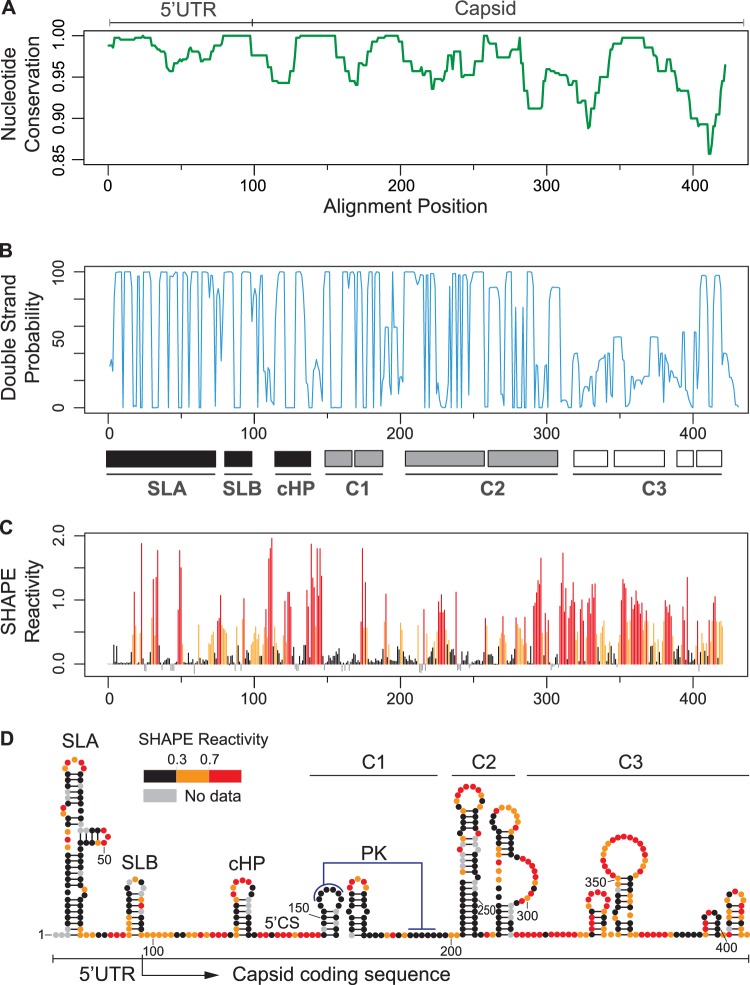
Conserved RNA structures in the DENV capsid coding sequence. (A) Nucleotide conservation of the first 450 nucleotides of the viral genome using representative sequences of the six different DENV2 genotypes. Window size = 10. (B) Double-stranded probability of the first 450 nucleotides of the viral genome. Three regions were defined: high double-strand probability for 5′ SLA, SLB, and cHP (black boxes); C1 and C2 structures (gray boxes); and unstructured C3 region (white boxes). (C) Plot showing SHAPE reactivity at each nucleotide position of DENV2 RNA. (D) Viral RNA structures based on conservation, predictions, and SHAPE reactivity. Red notations correspond to unpaired nucleotides, and black indicates double-stranded regions, whereas residues of intermediate reactivity are noted in orange. Abbreviations: UTR, untranslated region; SLA, stem-loop A; SLB, stem-loop B; cHP, capsid hairpin; 5′ CS, 5′ cyclization sequence; PK, pseudoknot; C1, C2, and C3, RNA structures identified in the capsid coding sequence. Data shown are averages of three experiments.
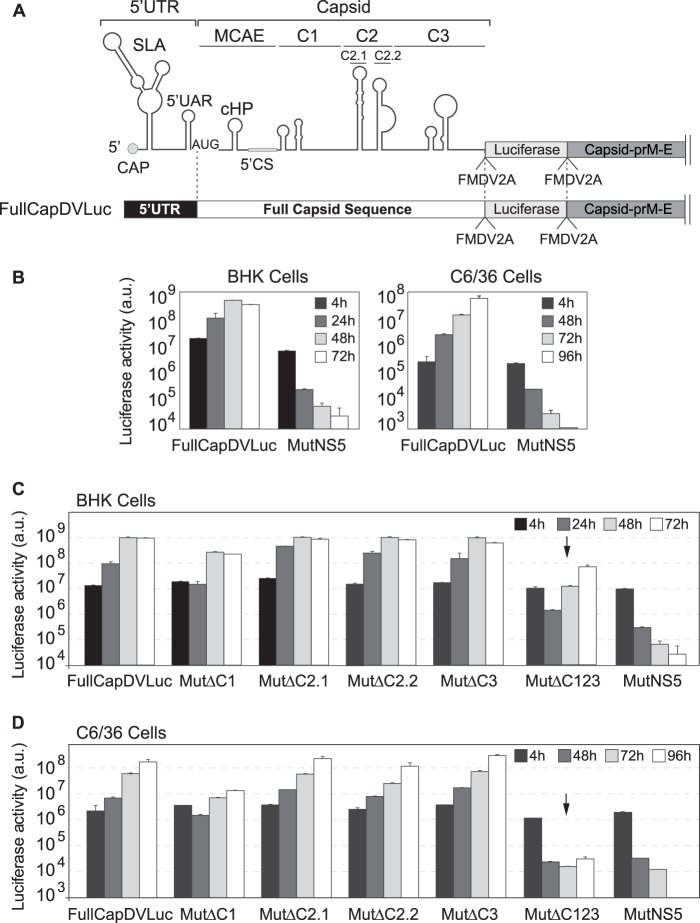
Functional significance of RNA structures in the capsid coding sequence. (A) Schematic representation of the DENV reporter FullCapDVLuc. FullCap and MCAE stand for full capsid coding sequence and minimal cis-acting elements, respectively. (B) Replication of new DENV reporter construct. Plots show Renilla luciferase activity as a function of time post-RNA transfection in BHK and C6/36 cells. The luciferase values are means ± standard deviations (n = 4). (C and D) Translation and replication of DENV reporter RNAs containing the indicated deletions in BHK cells (C) and C6/36 cells (D). Luciferase activity was measured as a function of time and is represented as arbitrary units (a.u.). The luciferase values are means ± standard deviations (n = 4).
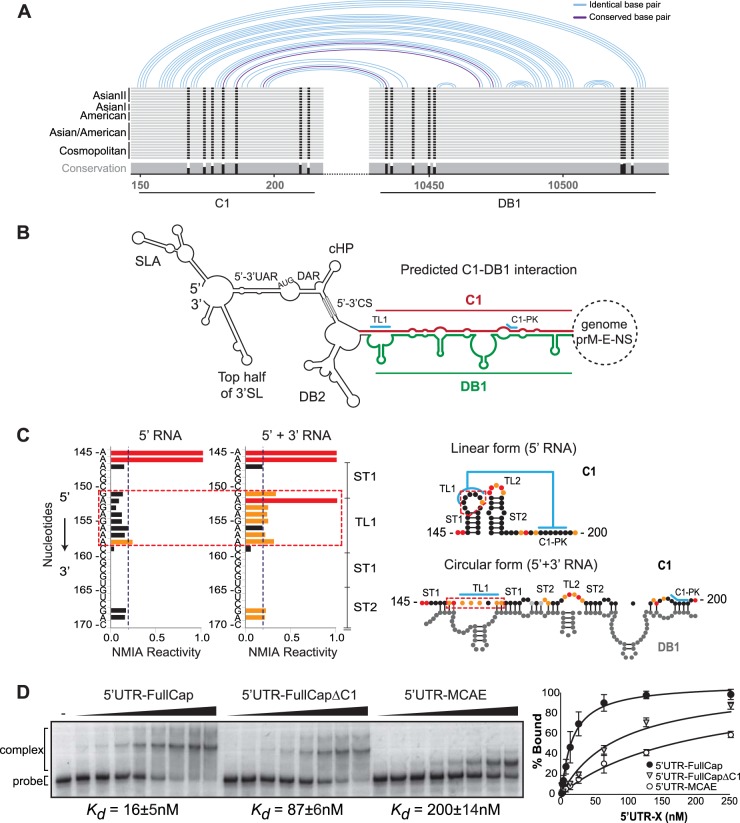
Sequence in the capsid protein coding region facilitates long-range RNA-RNA interactions between the ends of the viral genome. (A) Schematic representation of the conserved patterns of long-range interactions based on alignments of genomes of different DENV2 genotypes. Details of nucleotide sequences, alignments, and predictions are described in Fig. S1 in the supplemental material. (B) Representation of the circular conformation of the DENV genome indicating the known and new complementary elements. The predicted RNA-RNA interaction between C1 (red) and DB1 (green) is indicated. (C) Analysis of 5′-end RNA secondary structure in the absence and presence of the 3′ UTR RNA (ratio of 1:5). NMIA probing profiles of an RNA corresponding to the 5′ end of the genome (5′ RNA) and the probing of the same RNA in the presence of a second RNA containing the viral 3′ UTR sequence (5′ + 3′ RNA) are shown on the left. The y axis contains the sequence and nucleotide numbers; the x axis depicts chemical reactivity. On the right, secondary structures and NMIA reactivity are indicated for the predicted 5′ end alone and the 5′-3′ hybridized form. Red dashed frames highlight a region with different probing profiles. (D) Mobility shift assays indicate that C1-DB1 hybridization stabilizes the RNA-RNA complex. The 3′ UTR RNA probe was incubated with increasing concentrations of unlabeled RNAs corresponding to molecules with viral 5′-end sequences as indicated. Mobilities of the 3′ UTR RNA probe and RNA-RNA complex as well as the apparent dissociation constants (Kd ± standard error) are indicated. On the right, the percentage of RNA probe bound was plotted as a function of unlabeled RNA concentration for the three molecules used.
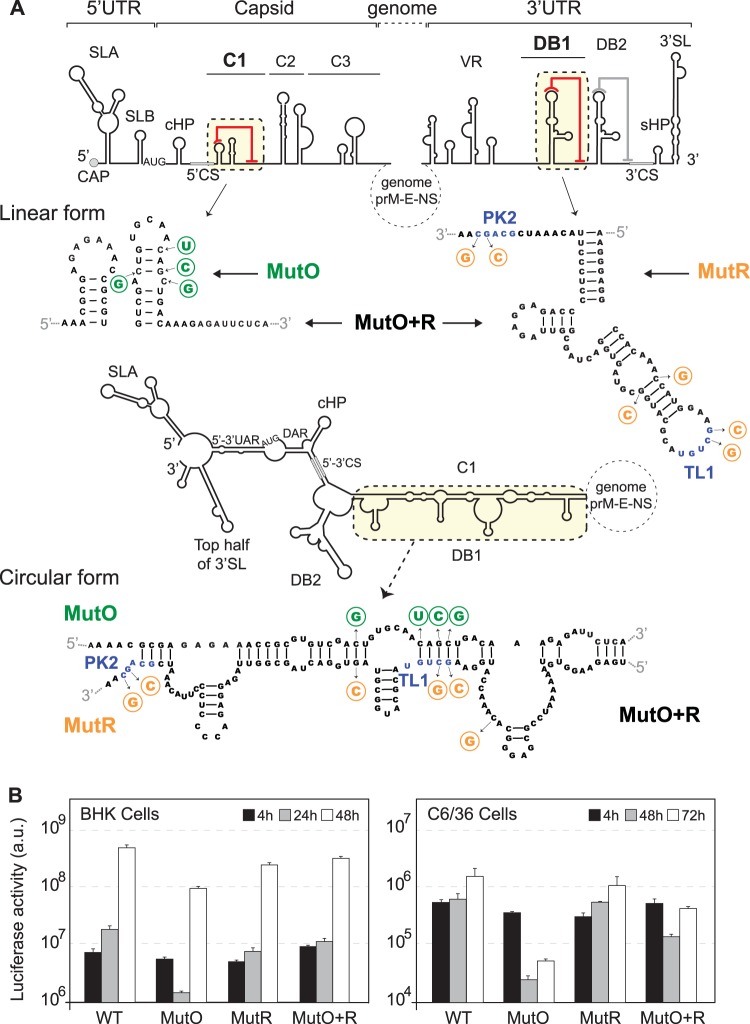
Relevance of C1-DB1 interaction in DENV RNA replication. (A) Design of mutation disrupting and reconstituting long-range C1-DB1 RNA-RNA interaction in the context of the viral genome. cis-acting elements in the 5′ end and the 3′ end of the genome are indicated. Nucleotide sequences of C1 and DB1 and the substitutions incorporated are shown in the two alternative conformations, linear and circular. Location, nucleotide changes, and names of mutants are indicated. MutO (green) shows substitutions in C1, and MutR (orange) shows substitutions in DB1. MutO+R contains both substitutions, restoring the interaction and maintaining C1 and DB1 structures. In addition, this mutant maintains the pseudoknot TL1/PK2 by compensatory mutations. (B) Replication of the three mutant RNAs in BHK and C6/36 cells together with the WT control. Luciferase activity was measured as a function of time and represented as arbitrary units (a.u.). The luciferase values are means ± standard deviations (n = 3).
Similar articles
-
Novel cis-acting element within the capsid-coding region enhances flavivirus viral-RNA replication by regulating genome cyclization.J Virol. 2013 Jun;87(12):6804-18. doi: 10.1128/JVI.00243-13. Epub 2013 Apr 10. J Virol. 2013. PMID: 23576500 Free PMC article.
-
RNA Structure Duplication in the Dengue Virus 3' UTR: Redundancy or Host Specificity?mBio. 2019 Jan 8;10(1):e02506-18. doi: 10.1128/mBio.02506-18. mBio. 2019. PMID: 30622191 Free PMC article.
-
Uncoupling cis-Acting RNA elements from coding sequences revealed a requirement of the N-terminal region of dengue virus capsid protein in virus particle formation.J Virol. 2012 Jan;86(2):1046-58. doi: 10.1128/JVI.05431-11. Epub 2011 Nov 9. J Virol. 2012. PMID: 22072762 Free PMC article.
-
Dynamic RNA structures in the dengue virus genome.RNA Biol. 2011 Mar-Apr;8(2):249-57. doi: 10.4161/rna.8.2.14992. Epub 2011 Mar 1. RNA Biol. 2011. PMID: 21593583 Review.
-
Functional RNA elements in the dengue virus genome.Viruses. 2011 Sep;3(9):1739-56. doi: 10.3390/v3091739. Epub 2011 Sep 15. Viruses. 2011. PMID: 21994804 Free PMC article. Review.
Cited by
-
The tale of two flaviviruses: subversion of host pathways by RNA shapes in dengue and hepatitis C viral RNA genomes.Curr Opin Microbiol. 2021 Feb;59:79-85. doi: 10.1016/j.mib.2020.08.007. Epub 2020 Oct 16. Curr Opin Microbiol. 2021. PMID: 33070015 Free PMC article. Review.
-
Evolutionary traits of Tick-borne encephalitis virus: Pervasive non-coding RNA structure conservation and molecular epidemiology.Virus Evol. 2022 Jun 11;8(1):veac051. doi: 10.1093/ve/veac051. eCollection 2022. Virus Evol. 2022. PMID: 35822110 Free PMC article.
-
Effects of Refolding on Large-Scale RNA Structure.Biochemistry. 2019 Jul 16;58(28):3069-3077. doi: 10.1021/acs.biochem.8b01219. Epub 2019 Jul 3. Biochemistry. 2019. PMID: 31268687 Free PMC article.
-
Flavors of Flaviviral RNA Structure: towards an Integrated View of RNA Function from Translation through Encapsidation.Bioessays. 2019 Aug;41(8):e1900003. doi: 10.1002/bies.201900003. Epub 2019 Jun 18. Bioessays. 2019. PMID: 31210384 Free PMC article. Review.
-
Genomic variations in the 3'-termini of Rice stripe virus in the rotation between vector insect and host plant.New Phytol. 2018 Aug;219(3):1085-1096. doi: 10.1111/nph.15246. Epub 2018 Jun 8. New Phytol. 2018. PMID: 29882354 Free PMC article.
References
-
- Gamarnik AV. 2010. Role of the dengue virus 5′ and 3′ untranslated regions in viral replication, p 55–78 InHanley KA, Weaver SC (ed), Frontiers in dengue virus research. Caister Academic Press, Wymondham, Norfolk, United Kingdom.
-
- Lindenbach BD, Thiel HJ, Rice CD. 2007. Flaviviridae: the viruses and their replication, p 1101–1152 InKnipe DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, Straus SE (ed), Fields virology, 5th ed, vol 1 Lippincott Williams & Wilkins, Philadelphia, PA.
-
- Alcaraz-Estrada SL, Yocupicio-Monroy M, del Angel RM. 2010. Insights into dengue virus genome replication. Future Virol 5:575–592. doi:10.2217/fvl.10.49. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
