

Pyruvate kinase and aspartate-glutamate carrier distributions reveal key metabolic links between neurons and glia in retina
- PMID: 25313047
- PMCID: PMC4217417
- DOI: 10.1073/pnas.1412441111
Pyruvate kinase and aspartate-glutamate carrier distributions reveal key metabolic links between neurons and glia in retina
Abstract
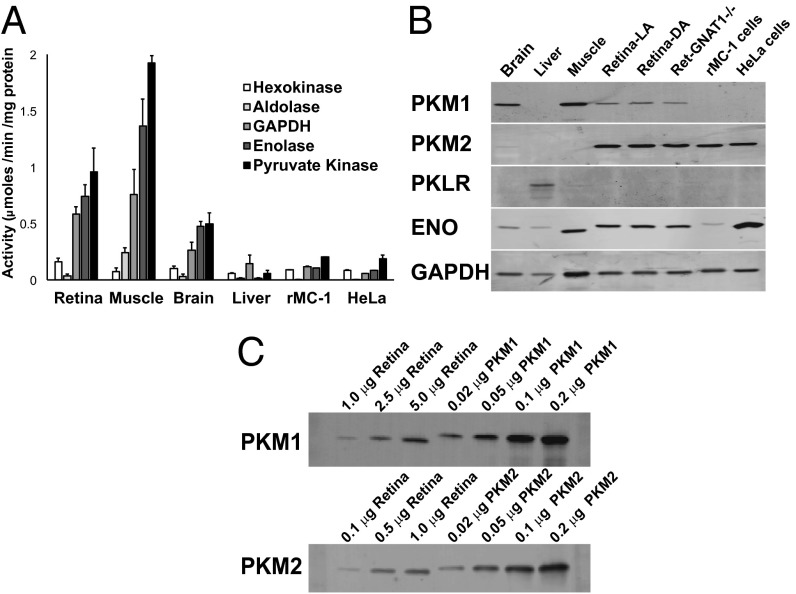
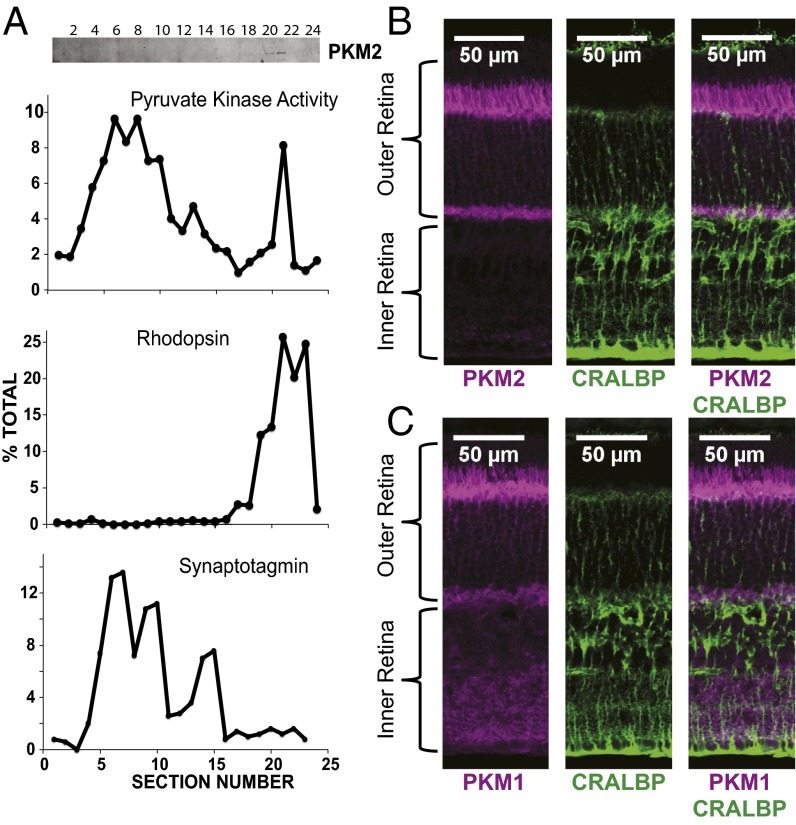
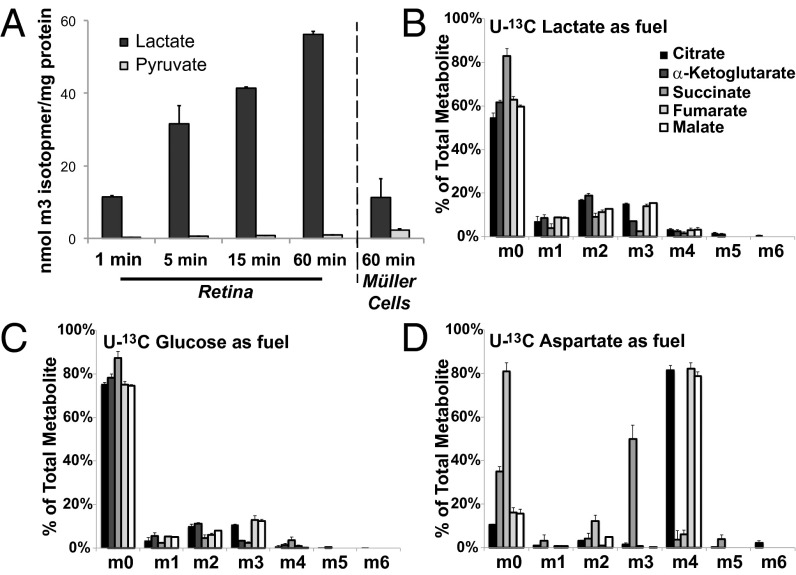
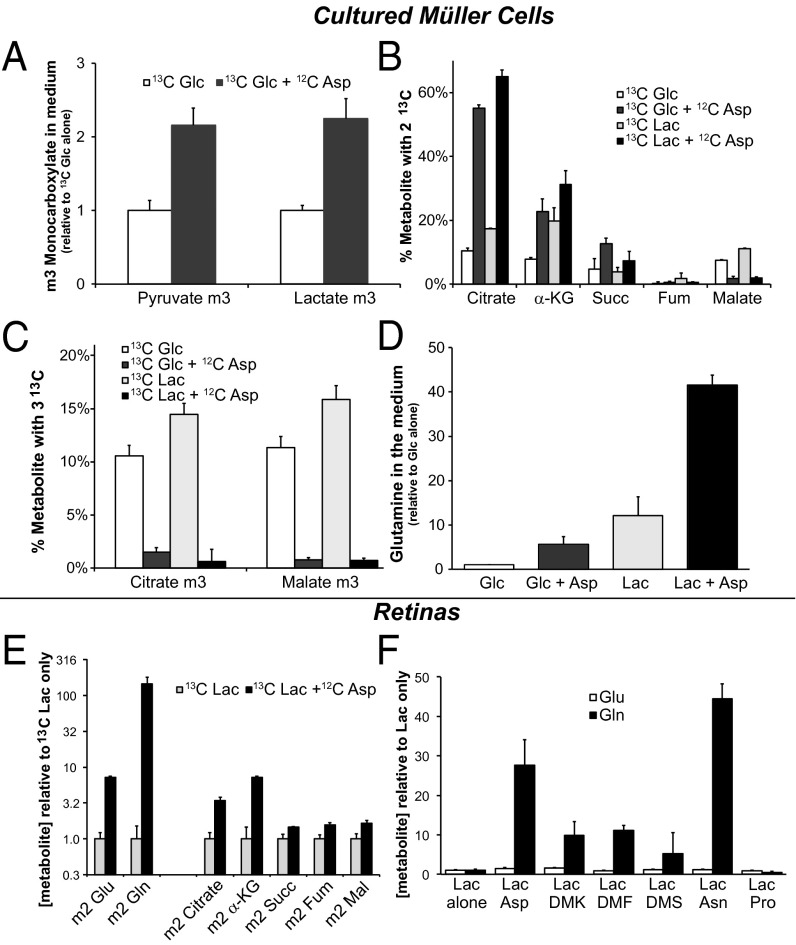
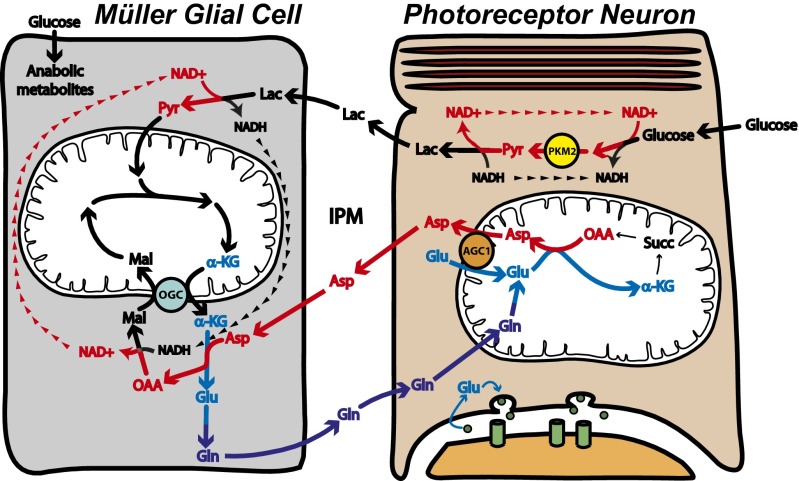
Symbiotic relationships between neurons and glia must adapt to structures, functions, and metabolic roles of the tissues they are in. We show here that Müller glia in retinas have specific enzyme deficiencies that can enhance their ability to synthesize Gln. The metabolic cost of these deficiencies is that they impair the Müller cell's ability to metabolize Glc. We show here that the cells can compensate for this deficiency by using metabolites produced by neurons. Müller glia are deficient for pyruvate kinase (PK) and for aspartate/glutamate carrier 1 (AGC1), a key component of the malate-aspartate shuttle. In contrast, photoreceptor neurons express AGC1 and the M2 isoform of pyruvate kinase, which is commonly associated with aerobic glycolysis in tumors, proliferating cells, and some other cell types. Our findings reveal a previously unidentified type of metabolic relationship between neurons and glia. Müller glia compensate for their unique metabolic adaptations by using lactate and aspartate from neurons as surrogates for their missing PK and AGC1.
Keywords: Müller glia; aerobic glycolysis; glutamine metabolism; photoreceptors; retina.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Molecular basis for increased lactate formation in the Müller glial cells of retina.Brain Res Bull. 2019 Jan;144:158-163. doi: 10.1016/j.brainresbull.2018.11.023. Epub 2018 Nov 29. Brain Res Bull. 2019. PMID: 30503222
-
Energy sources for glutamate neurotransmission in the retina: absence of the aspartate/glutamate carrier produces reliance on glycolysis in glia.J Neurochem. 2007 Apr;101(1):120-31. doi: 10.1111/j.1471-4159.2006.04349.x. J Neurochem. 2007. PMID: 17394462
-
AGC1 Deficiency: Pathology and Molecular and Cellular Mechanisms of the Disease.Int J Mol Sci. 2022 Jan 4;23(1):528. doi: 10.3390/ijms23010528. Int J Mol Sci. 2022. PMID: 35008954 Free PMC article. Review.
-
Deficient glucose and glutamine metabolism in Aralar/AGC1/Slc25a12 knockout mice contributes to altered visual function.Mol Vis. 2016 Oct 12;22:1198-1212. eCollection 2016. Mol Vis. 2016. PMID: 27746674 Free PMC article.
-
The mitochondrial aspartate/glutamate carrier AGC1 and calcium homeostasis: physiological links and abnormalities in autism.Mol Neurobiol. 2011 Aug;44(1):83-92. doi: 10.1007/s12035-011-8192-2. Epub 2011 Jun 21. Mol Neurobiol. 2011. PMID: 21691713 Review.
Cited by
-
Long-term HIF-1α stabilization reduces respiration, promotes mitophagy, and results in retinal cell death.Sci Rep. 2023 Nov 23;13(1):20541. doi: 10.1038/s41598-023-47942-8. Sci Rep. 2023. PMID: 37996657 Free PMC article.
-
Lactate: More Than Merely a Metabolic Waste Product in the Inner Retina.Mol Neurobiol. 2020 Apr;57(4):2021-2037. doi: 10.1007/s12035-019-01863-8. Epub 2020 Jan 8. Mol Neurobiol. 2020. PMID: 31916030 Review.
-
Higher Reliance on Glycolysis Limits Glycolytic Responsiveness in Degenerating Glaucomatous Optic Nerve.Mol Neurobiol. 2019 Oct;56(10):7097-7112. doi: 10.1007/s12035-019-1576-4. Epub 2019 Apr 13. Mol Neurobiol. 2019. PMID: 30980229 Free PMC article.
-
Biochemical Underpinnings of Immune Cell Metabolic Phenotypes.Immunity. 2017 May 16;46(5):703-713. doi: 10.1016/j.immuni.2017.04.013. Immunity. 2017. PMID: 28514672 Free PMC article. Review.
-
Probing Metabolism in the Intact Retina Using Stable Isotope Tracers.Methods Enzymol. 2015;561:149-70. doi: 10.1016/bs.mie.2015.04.002. Epub 2015 Jun 14. Methods Enzymol. 2015. PMID: 26358904 Free PMC article. Review.
References
-
- Gatenby RA, Gillies RJ. Why do cancers have high aerobic glycolysis? Nat Rev Cancer. 2004;4(11):891–899. - PubMed
-
- Lunt SY, Vander Heiden MG. Aerobic glycolysis: Meeting the metabolic requirements of cell proliferation. Annu Rev Cell Dev Biol. 2011;27:441–464. - PubMed
-
- Wang L, Kondo M, Bill A. Glucose metabolism in cat outer retina. Effects of light and hyperoxia. Invest Ophthalmol Vis Sci. 1997;38(1):48–55. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
