

Progressive replacement of embryo-derived cardiac macrophages with age
- PMID: 25245760
- PMCID: PMC4203946
- DOI: 10.1084/jem.20140639
Progressive replacement of embryo-derived cardiac macrophages with age
Abstract
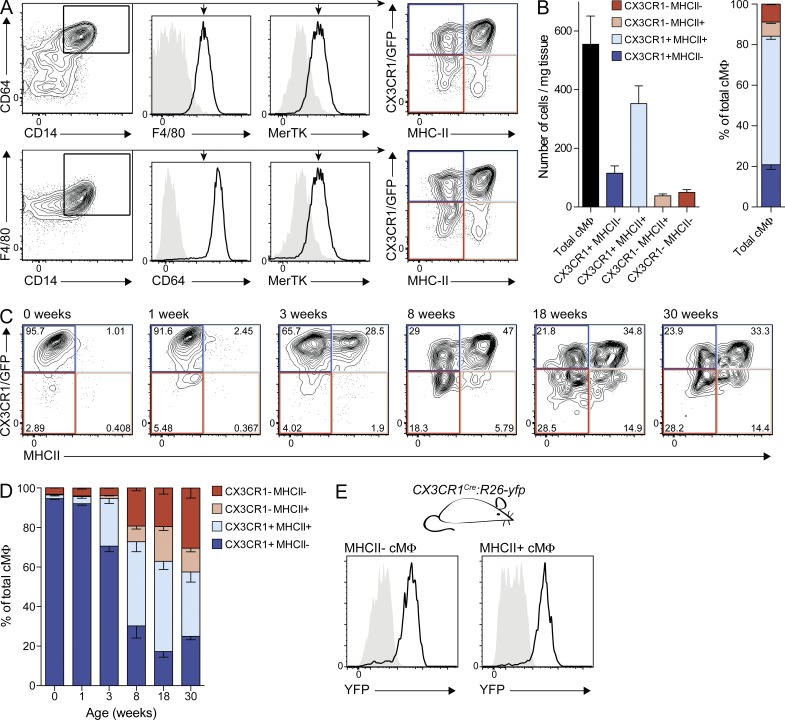
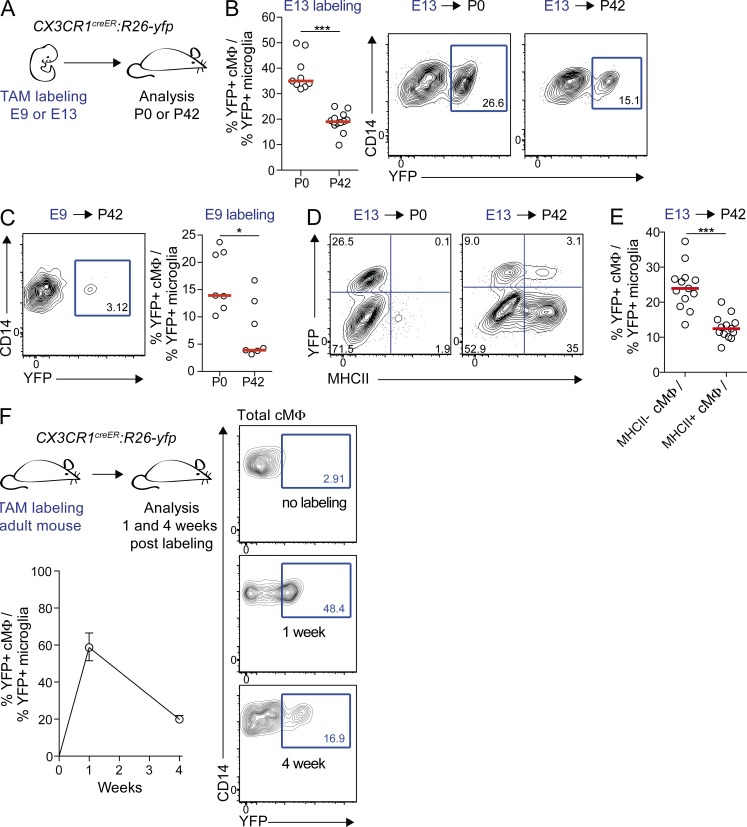
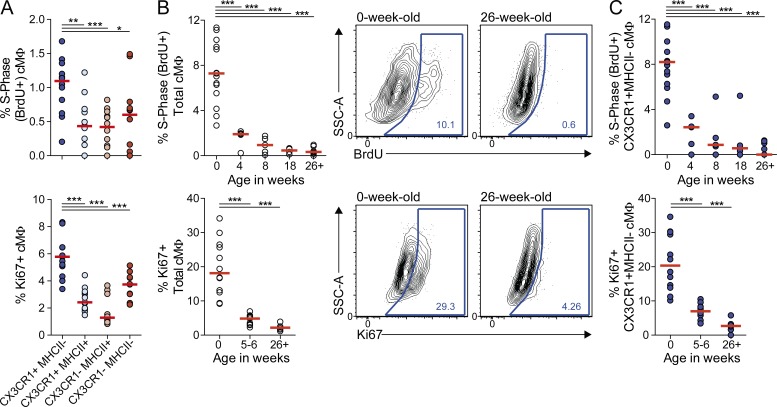
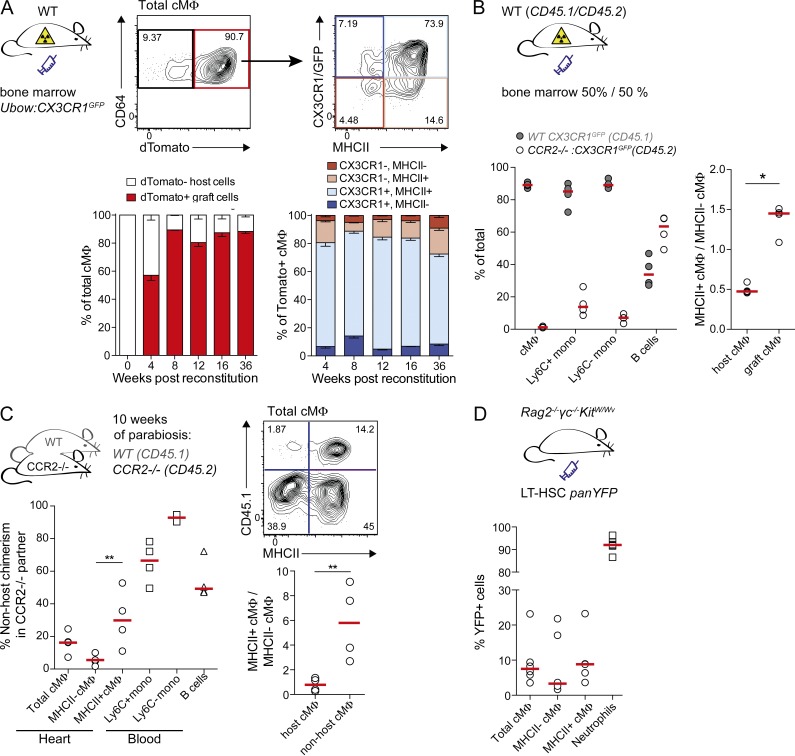
Cardiac macrophages (cMΦ) are critical for early postnatal heart regeneration and fibrotic repair in the adult heart, but their origins and cellular dynamics during postnatal development have not been well characterized. Tissue macrophages can be derived from embryonic progenitors or from monocytes during inflammation. We report that within the first weeks after birth, the embryo-derived population of resident CX3CR1(+) cMΦ diversifies into MHCII(+) and MHCII(-) cells. Genetic fate mapping demonstrated that cMΦ derived from CX3CR1(+) embryonic progenitors persisted into adulthood but the initially high contribution to resident cMΦ declined after birth. Consistent with this, the early significant proliferation rate of resident cMΦ decreased with age upon diversification into subpopulations. Bone marrow (BM) reconstitution experiments showed monocyte-dependent quantitative replacement of all cMΦ populations. Furthermore, parabiotic mice and BM chimeras of nonirradiated recipient mice revealed a slow but significant donor contribution to cMΦ. Together, our observations indicate that in the heart, embryo-derived cMΦ show declining self-renewal with age and are progressively substituted by monocyte-derived macrophages, even in the absence of inflammation.
© 2014 Molawi et al.
Figures
Comment in
-
Monocytes find a new place to dwell in the niche of heartbreak hotel.J Exp Med. 2014 Oct 20;211(11):2136. doi: 10.1084/jem.21111insight1. J Exp Med. 2014. PMID: 25332351 Free PMC article. No abstract available.
Similar articles
-
Self-renewing resident arterial macrophages arise from embryonic CX3CR1(+) precursors and circulating monocytes immediately after birth.Nat Immunol. 2016 Feb;17(2):159-68. doi: 10.1038/ni.3343. Epub 2015 Dec 7. Nat Immunol. 2016. PMID: 26642357
-
Bone marrow cells transdifferentiate to cardiomyocytes when introduced into the embryonic heart.Stem Cells. 2006 May;24(5):1236-45. doi: 10.1634/stemcells.2005-0128. Epub 2006 Jan 12. Stem Cells. 2006. PMID: 16410395
-
Embryonic and adult-derived resident cardiac macrophages are maintained through distinct mechanisms at steady state and during inflammation.Immunity. 2014 Jan 16;40(1):91-104. doi: 10.1016/j.immuni.2013.11.019. Immunity. 2014. PMID: 24439267 Free PMC article.
-
Tissue macrophage identity and self-renewal.Immunol Rev. 2014 Nov;262(1):56-73. doi: 10.1111/imr.12224. Immunol Rev. 2014. PMID: 25319327 Review.
-
Monocyte Heterogeneity: Consequences for Monocyte-Derived Immune Cells.J Immunol Res. 2016;2016:1475435. doi: 10.1155/2016/1475435. Epub 2016 Jul 11. J Immunol Res. 2016. PMID: 27478854 Free PMC article. Review.
Cited by
-
Neuroimmune interactions and osteoarthritis pain: focus on macrophages.Pain Rep. 2021 Mar 9;6(1):e892. doi: 10.1097/PR9.0000000000000892. eCollection 2021. Pain Rep. 2021. PMID: 33981927 Free PMC article. Review.
-
Aging dysregulates D- and E-series resolvins to modulate cardiosplenic and cardiorenal network following myocardial infarction.Aging (Albany NY). 2016 Oct 18;8(11):2611-2634. doi: 10.18632/aging.101077. Aging (Albany NY). 2016. PMID: 27777380 Free PMC article.
-
Using High-Dimensional Approaches to Probe Monocytes and Macrophages in Cardiovascular Disease.Front Immunol. 2019 Sep 12;10:2146. doi: 10.3389/fimmu.2019.02146. eCollection 2019. Front Immunol. 2019. PMID: 31572369 Free PMC article. Review.
-
Erythromyeloid progenitors give rise to a population of osteoclasts that contribute to bone homeostasis and repair.Nat Cell Biol. 2020 Jan;22(1):49-59. doi: 10.1038/s41556-019-0437-8. Epub 2020 Jan 6. Nat Cell Biol. 2020. PMID: 31907410 Free PMC article.
-
Early Development of Resident Macrophages in the Mouse Cochlea Depends on Yolk Sac Hematopoiesis.Front Neurol. 2019 Oct 22;10:1115. doi: 10.3389/fneur.2019.01115. eCollection 2019. Front Neurol. 2019. PMID: 31695671 Free PMC article.
References
-
- Chorro, L., Sarde A., Li M., Woollard K.J., Chambon P., Malissen B., Kissenpfennig A., Barbaroux J.-B., Groves R., and Geissmann F.. 2009. Langerhans cell (LC) proliferation mediates neonatal development, homeostasis, and inflammation-associated expansion of the epidermal LC network. J. Exp. Med. 206:3089–3100. 10.1084/jem.20091586 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
