

Commensal bacteria protect against food allergen sensitization
- PMID: 25157157
- PMCID: PMC4246970
- DOI: 10.1073/pnas.1412008111
Commensal bacteria protect against food allergen sensitization
Abstract
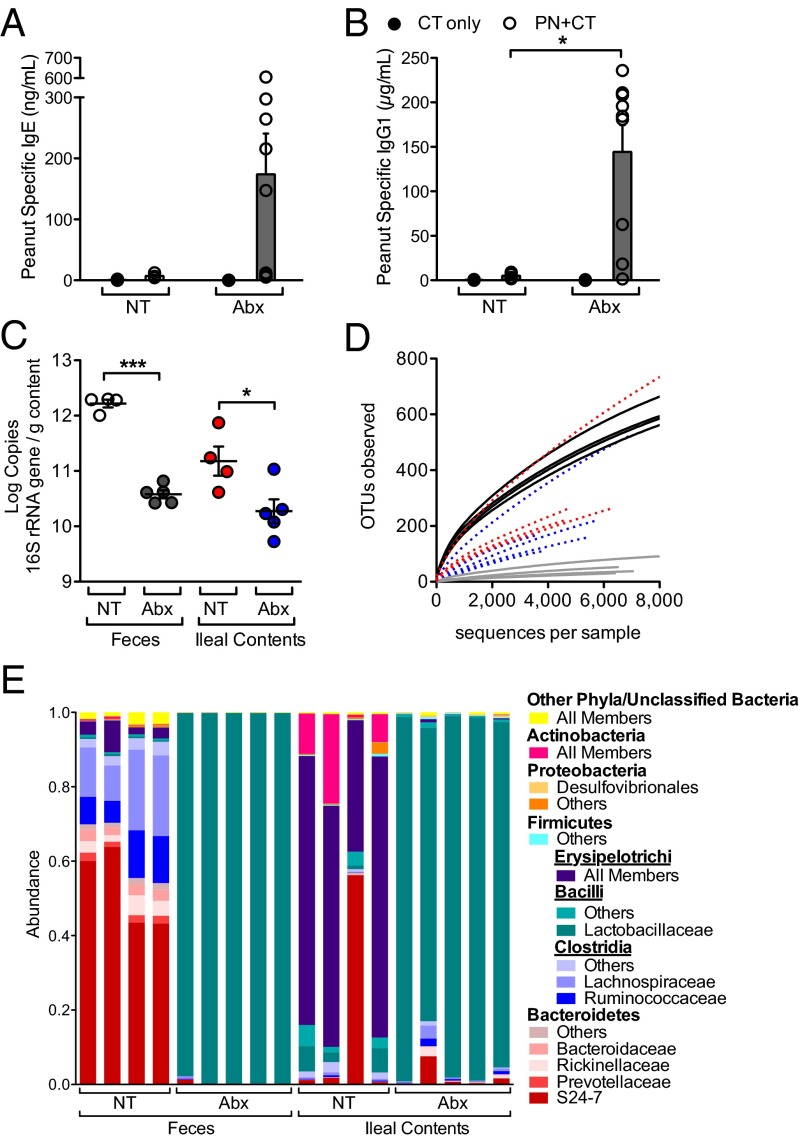
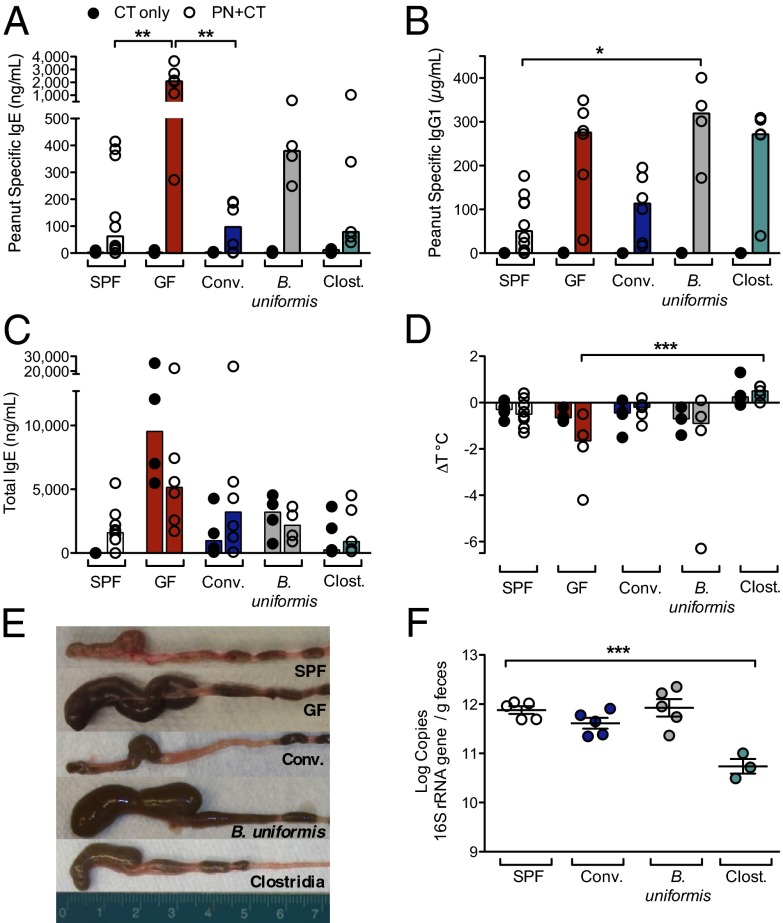
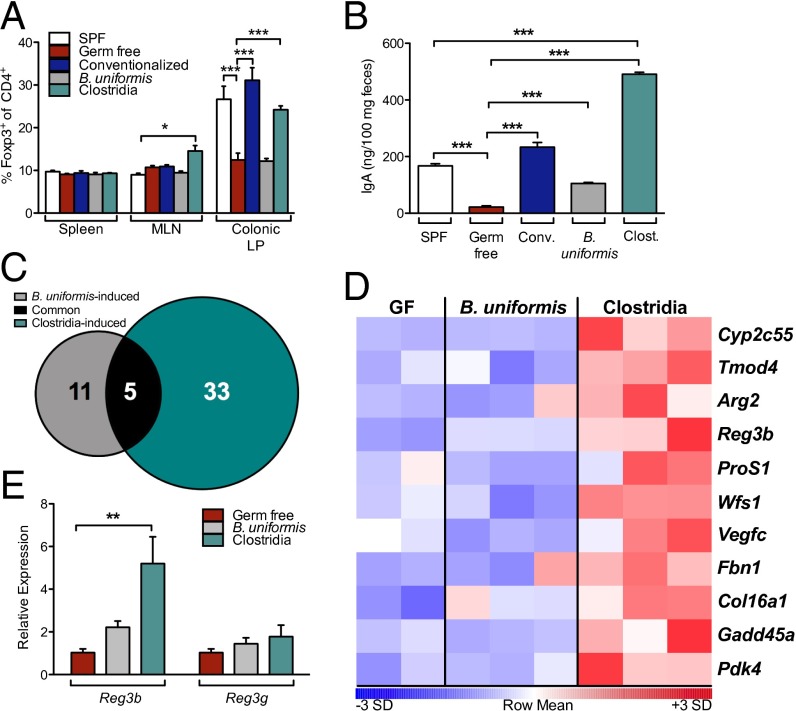
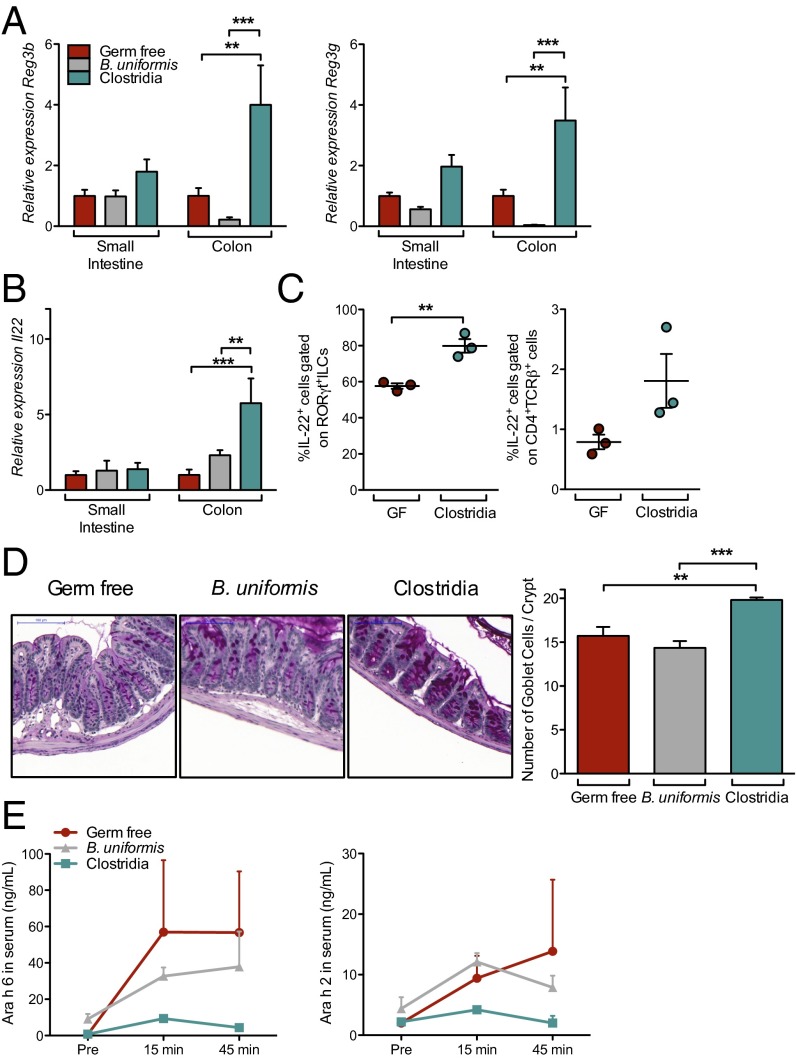
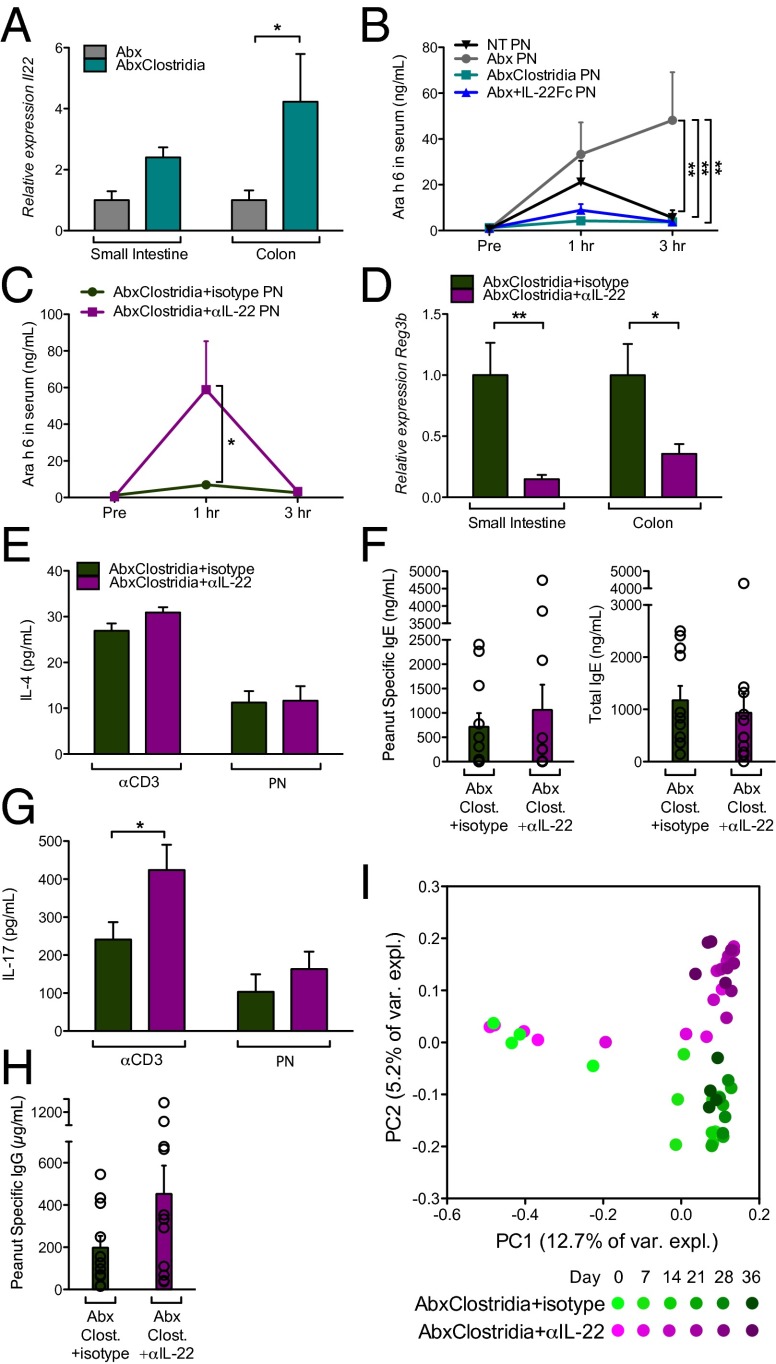
Environmentally induced alterations in the commensal microbiota have been implicated in the increasing prevalence of food allergy. We show here that sensitization to a food allergen is increased in mice that have been treated with antibiotics or are devoid of a commensal microbiota. By selectively colonizing gnotobiotic mice, we demonstrate that the allergy-protective capacity is conferred by a Clostridia-containing microbiota. Microarray analysis of intestinal epithelial cells from gnotobiotic mice revealed a previously unidentified mechanism by which Clostridia regulate innate lymphoid cell function and intestinal epithelial permeability to protect against allergen sensitization. Our findings will inform the development of novel approaches to prevent or treat food allergy based on modulating the composition of the intestinal microbiota.
Keywords: IL-22; barrier; microbiome.
Conflict of interest statement
Conflict of interest statement: A provisional US patent application (61/937952) was filed on February 10, 2014.
Figures
Similar articles
-
The role of the commensal microbiota in the regulation of tolerance to dietary allergens.Curr Opin Allergy Clin Immunol. 2015 Jun;15(3):243-9. doi: 10.1097/ACI.0000000000000157. Curr Opin Allergy Clin Immunol. 2015. PMID: 25827065 Free PMC article. Review.
-
The role of commensal bacteria in the regulation of sensitization to food allergens.FEBS Lett. 2014 Nov 17;588(22):4258-66. doi: 10.1016/j.febslet.2014.04.026. Epub 2014 May 1. FEBS Lett. 2014. PMID: 24791655 Free PMC article. Review.
-
Allergen Immunization Induces Major Changes in Microbiota Composition and Short-Chain Fatty Acid Production in Different Gut Segments in a Mouse Model of Lupine Food Allergy.Int Arch Allergy Immunol. 2018;177(4):311-323. doi: 10.1159/000492006. Epub 2018 Sep 21. Int Arch Allergy Immunol. 2018. PMID: 30244242
-
A microbiota signature associated with experimental food allergy promotes allergic sensitization and anaphylaxis.J Allergy Clin Immunol. 2013 Jan;131(1):201-12. doi: 10.1016/j.jaci.2012.10.026. Epub 2012 Nov 30. J Allergy Clin Immunol. 2013. PMID: 23201093 Free PMC article.
-
Prevention of birch pollen-related food allergy by mucosal treatment with multi-allergen-chimers in mice.PLoS One. 2012;7(6):e39409. doi: 10.1371/journal.pone.0039409. Epub 2012 Jun 29. PLoS One. 2012. PMID: 22768077 Free PMC article.
Cited by
-
Dominant immune tolerance in the intestinal tract imposed by RelB-dependent migratory dendritic cells regulates protective type 2 immunity.Nat Commun. 2024 Oct 23;15(1):9143. doi: 10.1038/s41467-024-53112-9. Nat Commun. 2024. PMID: 39443450 Free PMC article.
-
Gut microbiome impact on childhood allergic rhinitis and house dust mite IgE responses.Pediatr Res. 2024 Oct 21. doi: 10.1038/s41390-024-03645-y. Online ahead of print. Pediatr Res. 2024. PMID: 39433961
-
Species- and subspecies-level characterization of health-associated bacterial consortia that colonize the human gut during infancy.Gut Microbes. 2024 Jan-Dec;16(1):2414975. doi: 10.1080/19490976.2024.2414975. Epub 2024 Oct 20. Gut Microbes. 2024. PMID: 39428758 Free PMC article.
-
Oral microbiome in human health and diseases.mLife. 2024 Sep 16;3(3):367-383. doi: 10.1002/mlf2.12136. eCollection 2024 Sep. mLife. 2024. PMID: 39359681 Free PMC article. Review.
-
Tackling food allergies by microbiome modulation.Nat Mater. 2024 Oct;23(10):1318-1319. doi: 10.1038/s41563-024-02009-5. Nat Mater. 2024. PMID: 39349655 No abstract available.
References
-
- Wills-Karp M, Santeliz J, Karp CL. The germless theory of allergic disease: Revisiting the hygiene hypothesis. Nat Rev Immunol. 2001;1(1):69–75. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- UL1 TR000430/TR/NCATS NIH HHS/United States
- AI089954/AI/NIAID NIH HHS/United States
- P30 DK042086/DK/NIDDK NIH HHS/United States
- AI091962/AI/NIAID NIH HHS/United States
- T32 CA009547/CA/NCI NIH HHS/United States
- DK42086/DK/NIDDK NIH HHS/United States
- R37 DK047722/DK/NIDDK NIH HHS/United States
- T32 AI007090/AI/NIAID NIH HHS/United States
- R21 AI091962/AI/NIAID NIH HHS/United States
- R01 AI106302/AI/NIAID NIH HHS/United States
- AI106302/AI/NIAID NIH HHS/United States
- T32AI007090-33/AI/NIAID NIH HHS/United States
- R01 AI089954/AI/NIAID NIH HHS/United States
- R01 DK105562/DK/NIDDK NIH HHS/United States
- R01 DK047722/DK/NIDDK NIH HHS/United States
- DK078938/DK/NIDDK NIH HHS/United States
- R01 DK078938/DK/NIDDK NIH HHS/United States
- R56 DK078938/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
