

mTORC2-SGK-1 acts in two environmentally responsive pathways with opposing effects on longevity
- PMID: 25040785
- PMCID: PMC4172656
- DOI: 10.1111/acel.12248
mTORC2-SGK-1 acts in two environmentally responsive pathways with opposing effects on longevity
Abstract
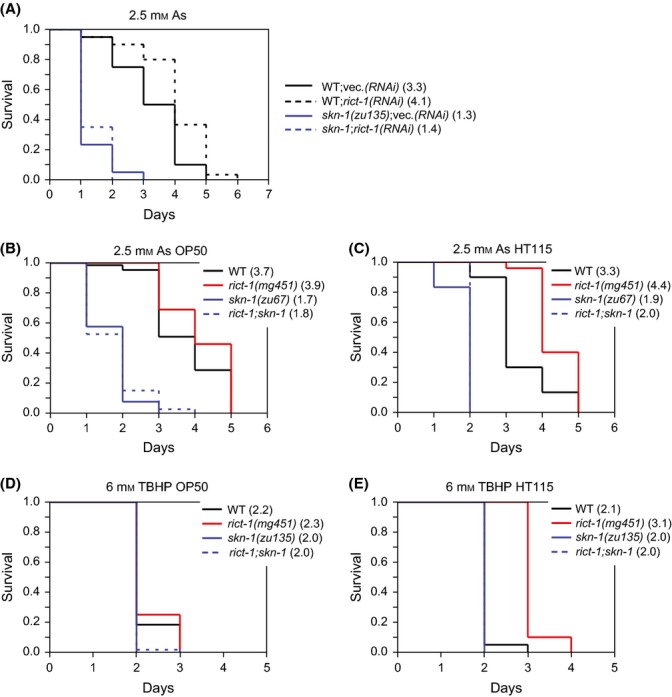
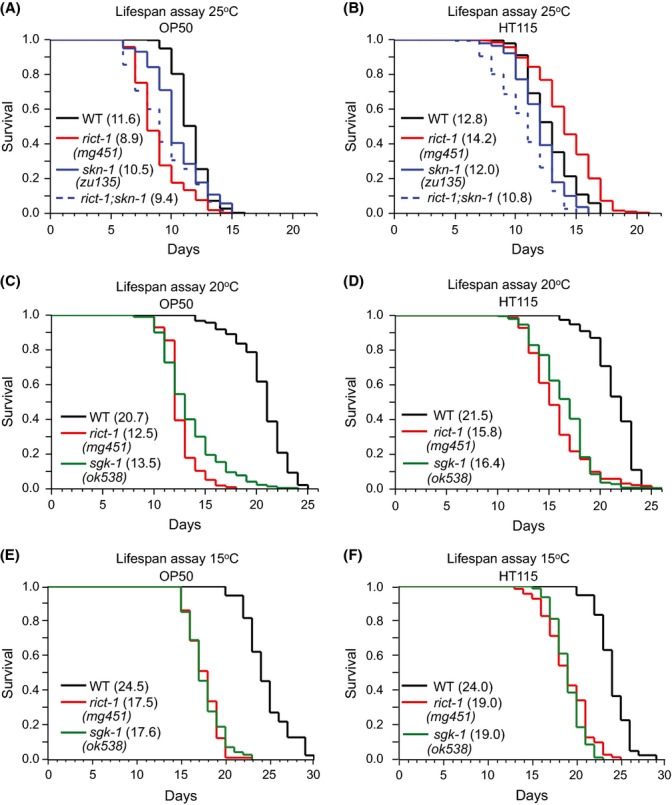
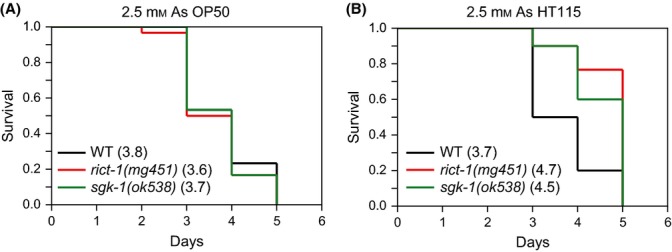
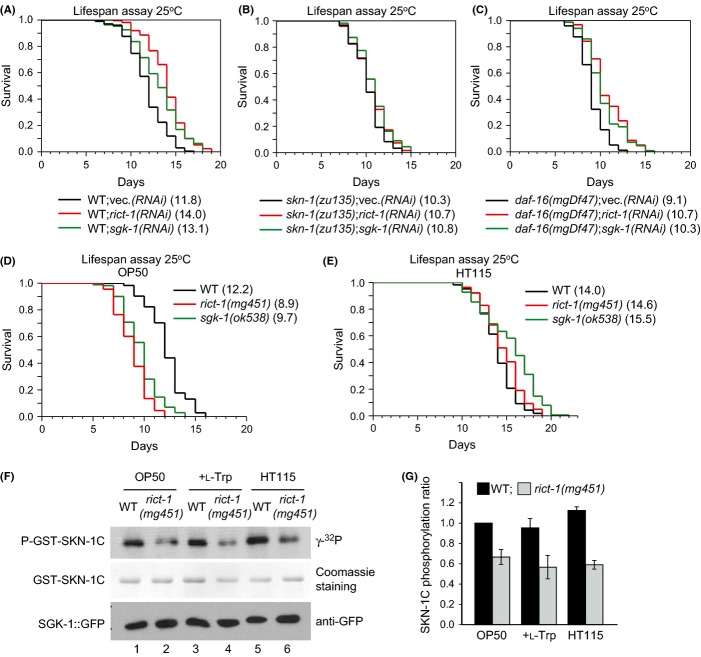
The nematode worm Caenorhabditis elegans provides a powerful system for elucidating how genetic, metabolic, nutritional, and environmental factors influence aging. The mechanistic target of rapamycin (mTOR) kinase is important in growth, disease, and aging and is present in the mTORC1 and mTORC2 complexes. In diverse eukaryotes, lifespan can be increased by inhibition of mTORC1, which transduces anabolic signals to stimulate protein synthesis and inhibit autophagy. Less is understood about mTORC2, which affects C. elegans lifespan in a complex manner that is influenced by the bacterial food source. mTORC2 regulates C. elegans growth, reproduction, and lipid metabolism by activating the SGK-1 kinase, but current data on SGK-1 and lifespan seem to be conflicting. Here, by analyzing the mTORC2 component Rictor (RICT-1), we show that mTORC2 modulates longevity by activating SGK-1 in two pathways that affect lifespan oppositely. RICT-1/mTORC2 limits longevity by directing SGK-1 to inhibit the stress-response transcription factor SKN-1/Nrf in the intestine. Signals produced by the bacterial food source determine how this pathway affects SKN-1 and lifespan. In addition, RICT-1/mTORC2 functions in neurons in an SGK-1-mediated pathway that increases lifespan at lower temperatures. RICT-1/mTORC2 and SGK-1 therefore oppose or accelerate aging depending upon the context in which they are active. Our findings reconcile data on SGK-1 and aging, show that the bacterial microenvironment influences SKN-1/Nrf, mTORC2 functions, and aging, and identify two longevity-related mTORC2 functions that involve SGK-regulated responses to environmental cues.
Keywords: SKN-1/Nrf; aging; mTORC2; microbiome; rictor; serum- and glucocorticoid-regulated kinase.
© 2014 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Figures
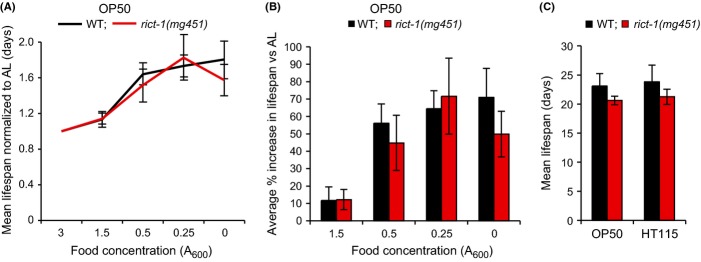
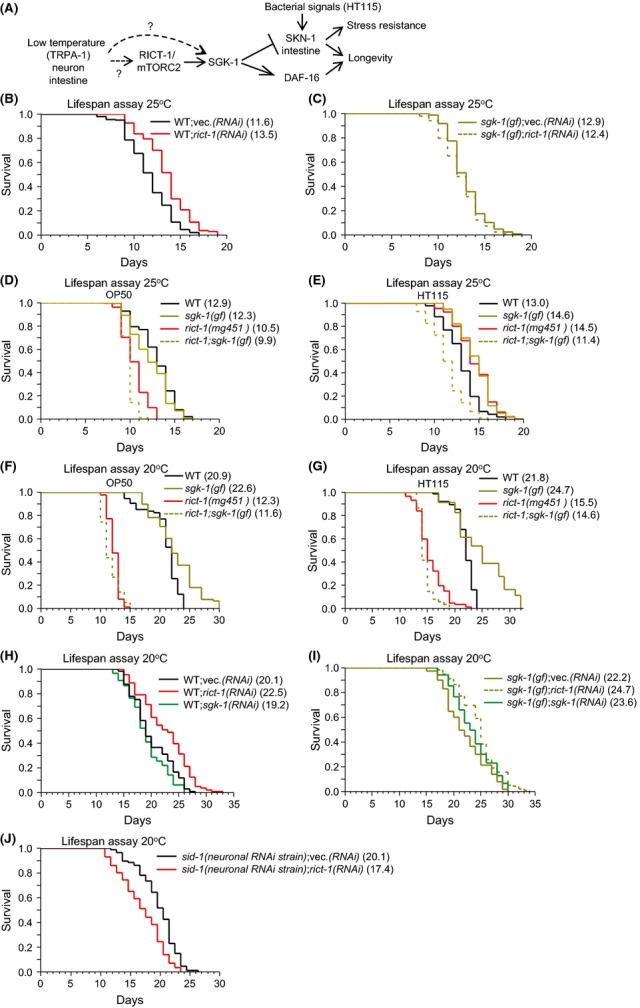
Similar articles
-
A non-canonical role for the C. elegans dosage compensation complex in growth and metabolic regulation downstream of TOR complex 2.Development. 2013 Sep;140(17):3601-12. doi: 10.1242/dev.094292. Epub 2013 Jul 24. Development. 2013. PMID: 23884442 Free PMC article.
-
TORC2 signaling antagonizes SKN-1 to induce C. elegans mesendodermal embryonic development.Dev Biol. 2013 Dec 15;384(2):214-27. doi: 10.1016/j.ydbio.2013.08.011. Epub 2013 Aug 20. Dev Biol. 2013. PMID: 23973804 Free PMC article.
-
Prohibitin-mediated lifespan and mitochondrial stress implicate SGK-1, insulin/IGF and mTORC2 in C. elegans.PLoS One. 2014 Sep 29;9(9):e107671. doi: 10.1371/journal.pone.0107671. eCollection 2014. PLoS One. 2014. PMID: 25265021 Free PMC article.
-
Growing knowledge of the mTOR signaling network.Semin Cell Dev Biol. 2014 Dec;36:79-90. doi: 10.1016/j.semcdb.2014.09.011. Epub 2014 Sep 19. Semin Cell Dev Biol. 2014. PMID: 25242279 Free PMC article. Review.
-
Targeted Inhibition of Rictor/mTORC2 in Cancer Treatment: A New Era after Rapamycin.Curr Cancer Drug Targets. 2016;16(4):288-304. doi: 10.2174/1568009616666151113120830. Curr Cancer Drug Targets. 2016. PMID: 26563881 Review.
Cited by
-
Inhibition of the Mechanistic Target of Rapamycin (mTOR)-Rapamycin and Beyond.Cold Spring Harb Perspect Med. 2016 May 2;6(5):a025924. doi: 10.1101/cshperspect.a025924. Cold Spring Harb Perspect Med. 2016. PMID: 27048303 Free PMC article. Review.
-
mTORC Inhibitors as Broad-Spectrum Therapeutics for Age-Related Diseases.Int J Mol Sci. 2018 Aug 8;19(8):2325. doi: 10.3390/ijms19082325. Int J Mol Sci. 2018. PMID: 30096787 Free PMC article. Review.
-
C. elegans DAF-16/FOXO interacts with TGF-ß/BMP signaling to induce germline tumor formation via mTORC1 activation.PLoS Genet. 2017 May 26;13(5):e1006801. doi: 10.1371/journal.pgen.1006801. eCollection 2017 May. PLoS Genet. 2017. PMID: 28549065 Free PMC article.
-
The Complex Roles of Mechanistic Target of Rapamycin in Adipocytes and Beyond.Trends Endocrinol Metab. 2017 May;28(5):319-339. doi: 10.1016/j.tem.2017.01.004. Epub 2017 Feb 22. Trends Endocrinol Metab. 2017. PMID: 28237819 Free PMC article. Review.
-
Tribbles pseudokinase NIPI-3 regulates intestinal immunity in Caenorhabditis elegans by controlling SKN-1/Nrf activity.Cell Rep. 2021 Aug 17;36(7):109529. doi: 10.1016/j.celrep.2021.109529. Cell Rep. 2021. PMID: 34407394 Free PMC article.
References
-
- Bowerman B, Eaton BA, Priess JR. skn-1, a maternally expressed gene required to specify the fate of ventral blastomeres in the early C. elegans embryo. Cell. 1992;68:1061–1075. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
