

Merlin/NF2 loss-driven tumorigenesis linked to CRL4(DCAF1)-mediated inhibition of the hippo pathway kinases Lats1 and 2 in the nucleus
- PMID: 25026211
- PMCID: PMC4126592
- DOI: 10.1016/j.ccr.2014.05.001
Merlin/NF2 loss-driven tumorigenesis linked to CRL4(DCAF1)-mediated inhibition of the hippo pathway kinases Lats1 and 2 in the nucleus
Abstract
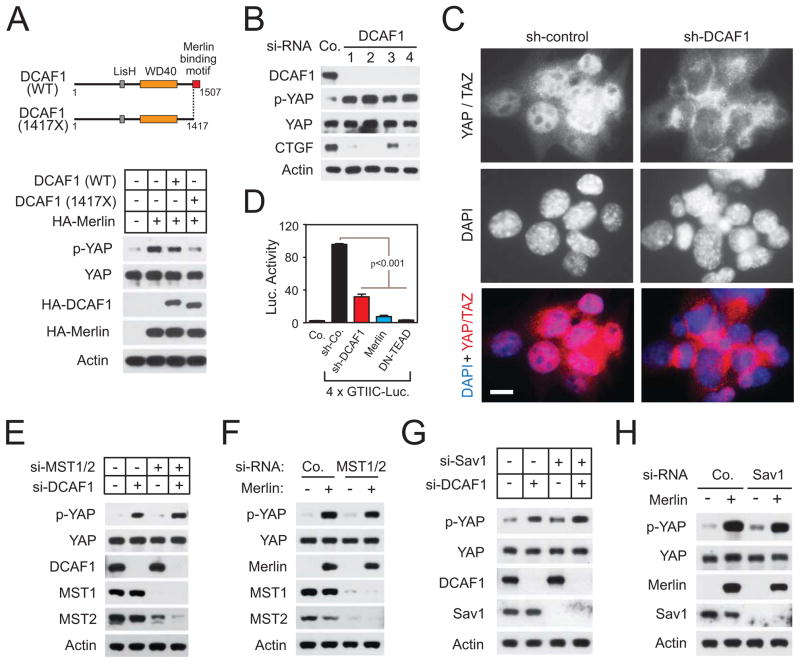
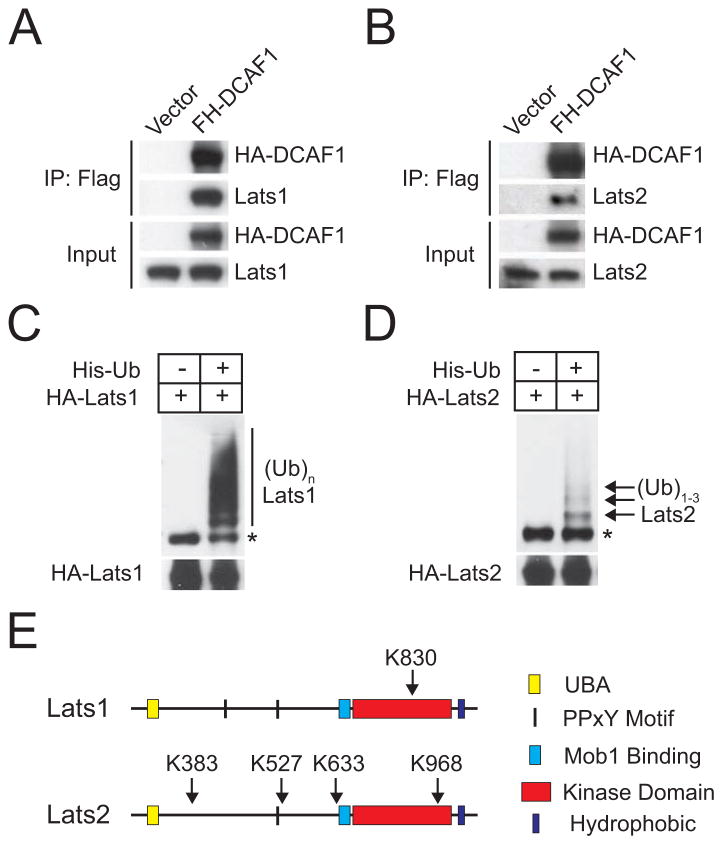
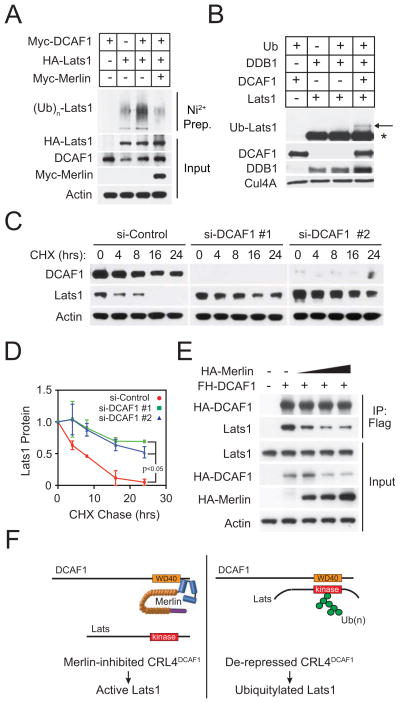
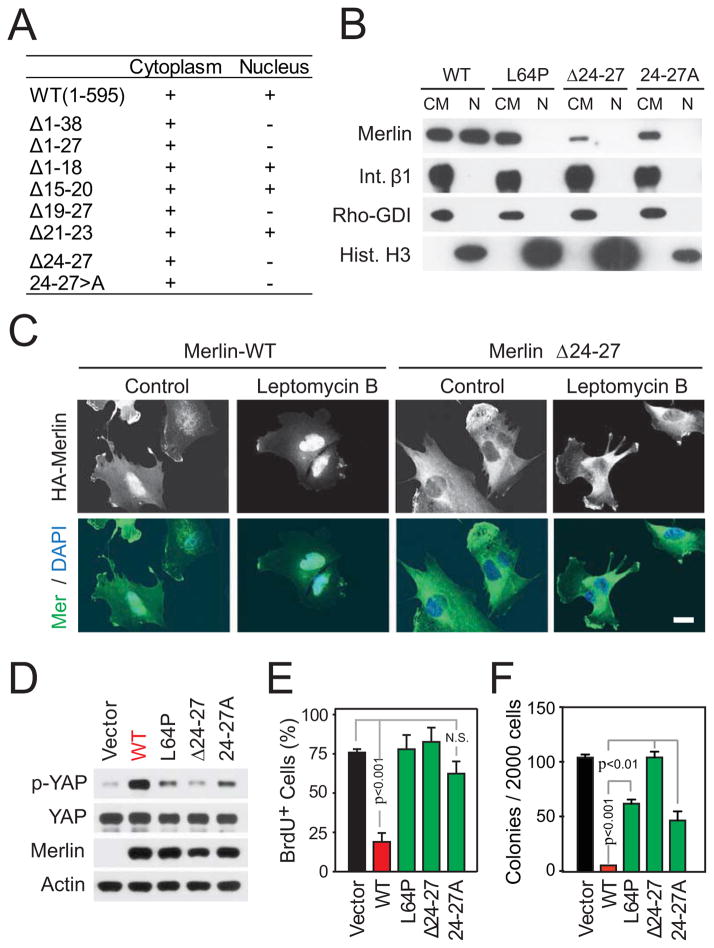
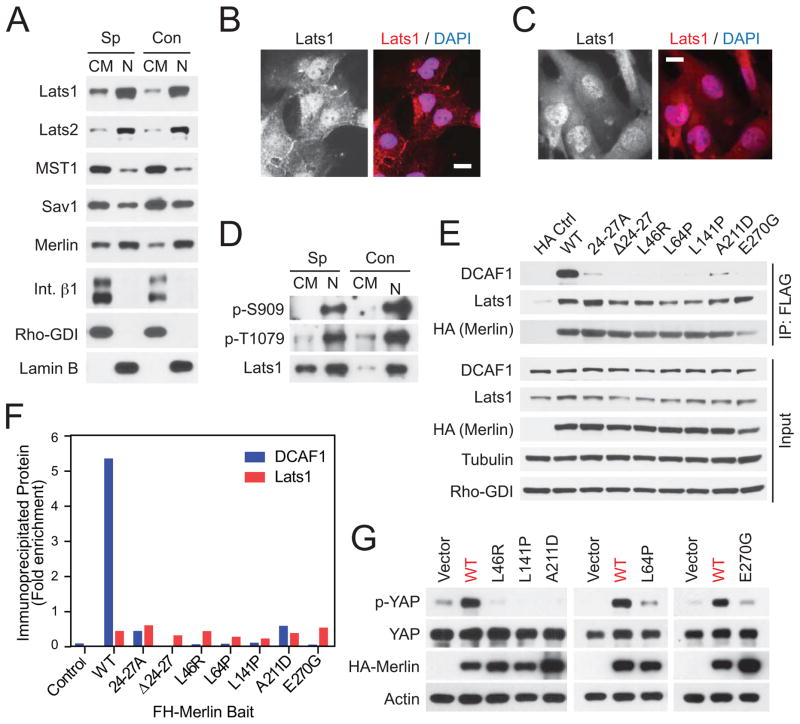
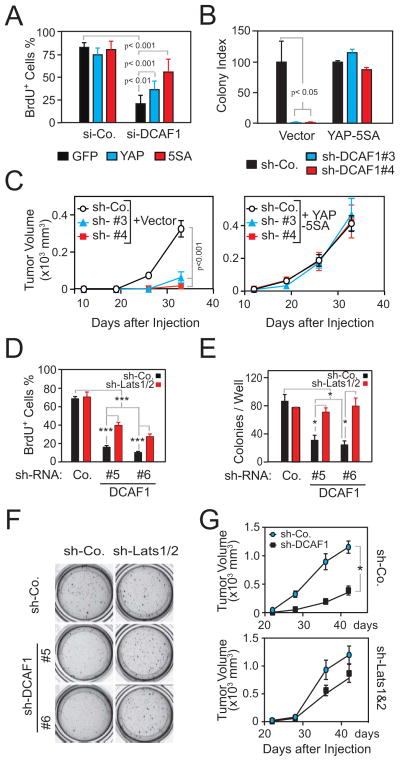
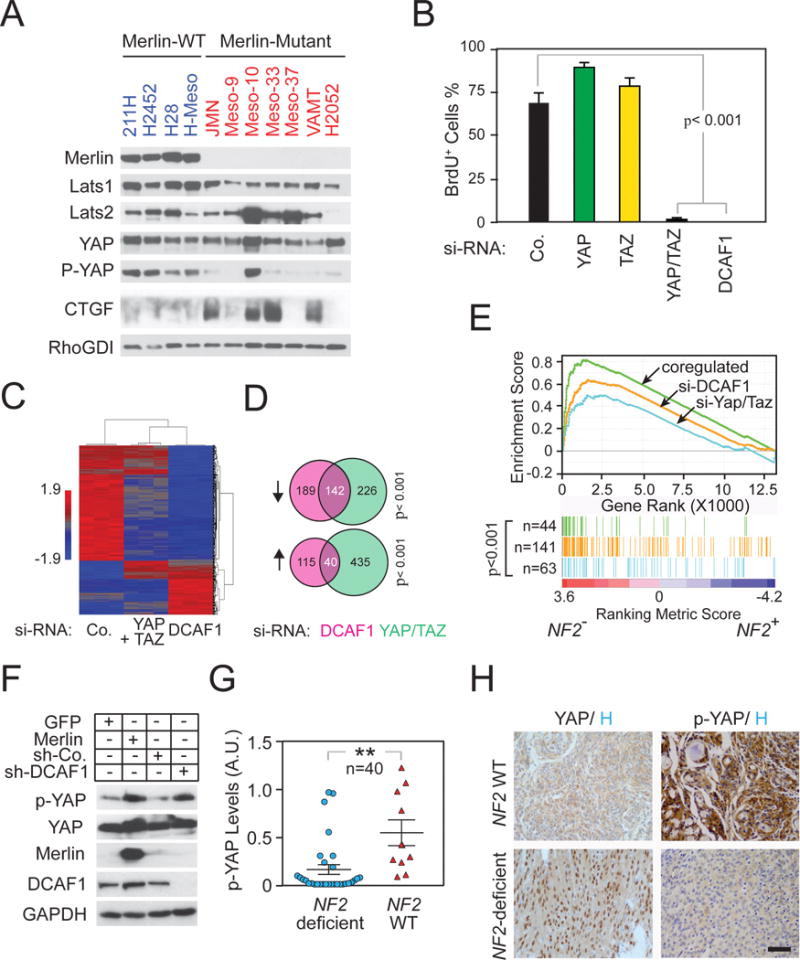
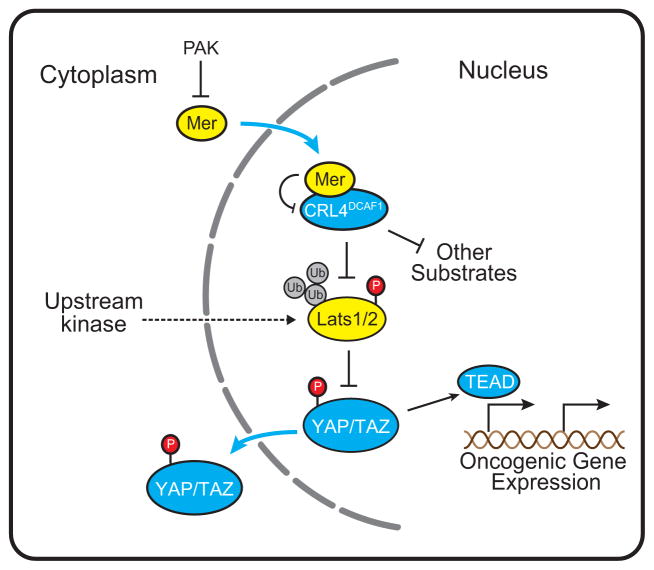
It is currently unclear whether Merlin/NF2 suppresses tumorigenesis by activating upstream components of the Hippo pathway at the plasma membrane or by inhibiting the E3 ubiquitin ligase CRL4(DCAF1) in the nucleus. We found that derepressed CRL4(DCAF1) promotes YAP- and TEAD-dependent transcription by ubiquitylating and, thereby, inhibiting Lats1 and 2 in the nucleus. Genetic epistasis experiments and analysis of tumor-derived missense mutations indicate that this signaling connection sustains the oncogenicity of Merlin-deficient tumor cells. Analysis of clinical samples confirms that this pathway operates in NF2-mutant tumors. We conclude that derepressed CRL4(DCAF1) promotes activation of YAP by inhibiting Lats1 and 2 in the nucleus.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Merlin/NF2 suppresses tumorigenesis by inhibiting the E3 ubiquitin ligase CRL4(DCAF1) in the nucleus.Cell. 2010 Feb 19;140(4):477-90. doi: 10.1016/j.cell.2010.01.029. Cell. 2010. PMID: 20178741 Free PMC article.
-
Merlin's tumor suppression linked to inhibition of the E3 ubiquitin ligase CRL4 (DCAF1).Cell Cycle. 2010 Nov 15;9(22):4433-6. doi: 10.4161/cc.9.22.13838. Epub 2010 Nov 15. Cell Cycle. 2010. PMID: 21084862 Free PMC article.
-
Statin suppresses Hippo pathway-inactivated malignant mesothelioma cells and blocks the YAP/CD44 growth stimulatory axis.Cancer Lett. 2017 Jan 28;385:215-224. doi: 10.1016/j.canlet.2016.10.020. Epub 2016 Oct 20. Cancer Lett. 2017. PMID: 27773750
-
Merlin, a multi-suppressor from cell membrane to the nucleus.FEBS Lett. 2012 May 21;586(10):1403-8. doi: 10.1016/j.febslet.2012.03.016. Epub 2012 Mar 21. FEBS Lett. 2012. PMID: 22595235 Review.
-
Molecular insights into NF2/Merlin tumor suppressor function.FEBS Lett. 2014 Aug 19;588(16):2743-52. doi: 10.1016/j.febslet.2014.04.001. Epub 2014 Apr 12. FEBS Lett. 2014. PMID: 24726726 Free PMC article. Review.
Cited by
-
New insights into posttranslational modifications of Hippo pathway in carcinogenesis and therapeutics.Cell Div. 2016 Mar 31;11:4. doi: 10.1186/s13008-016-0013-6. eCollection 2016. Cell Div. 2016. PMID: 27042197 Free PMC article. Review.
-
Disease implications of the Hippo/YAP pathway.Trends Mol Med. 2015 Apr;21(4):212-22. doi: 10.1016/j.molmed.2015.01.003. Epub 2015 Feb 18. Trends Mol Med. 2015. PMID: 25702974 Free PMC article. Review.
-
LATS1 but not LATS2 represses autophagy by a kinase-independent scaffold function.Nat Commun. 2019 Dec 17;10(1):5755. doi: 10.1038/s41467-019-13591-7. Nat Commun. 2019. PMID: 31848340 Free PMC article.
-
Induction of store-operated calcium entry (SOCE) suppresses glioblastoma growth by inhibiting the Hippo pathway transcriptional coactivators YAP/TAZ.Oncogene. 2019 Jan;38(1):120-139. doi: 10.1038/s41388-018-0425-7. Epub 2018 Aug 6. Oncogene. 2019. PMID: 30082911 Free PMC article.
-
Combined inhibition of Bcl-2 family members and YAP induces synthetic lethality in metastatic gastric cancer with RASA1 and NF2 deficiency.Mol Cancer. 2023 Sep 20;22(1):156. doi: 10.1186/s12943-023-01857-0. Mol Cancer. 2023. PMID: 37730636 Free PMC article.
References
-
- Angers S, Li T, Yi X, MacCoss MJ, Moon RT, Zheng N. Molecular architecture and assembly of the DDB1-CUL4A ubiquitin ligase machinery. Nature. 2006;443:590–593. - PubMed
-
- Chan EH, Nousiainen M, Chalamalasetty RB, Schafer A, Nigg EA, Sillje HH. The Ste20-like kinase Mst2 activates the human large tumor suppressor kinase Lats1. Oncogene. 2005;24:2076–2086. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
