

Regulation of germinal center responses and B-cell memory by the chromatin modifier MOZ
- PMID: 24979783
- PMCID: PMC4084455
- DOI: 10.1073/pnas.1402485111
Regulation of germinal center responses and B-cell memory by the chromatin modifier MOZ
Abstract
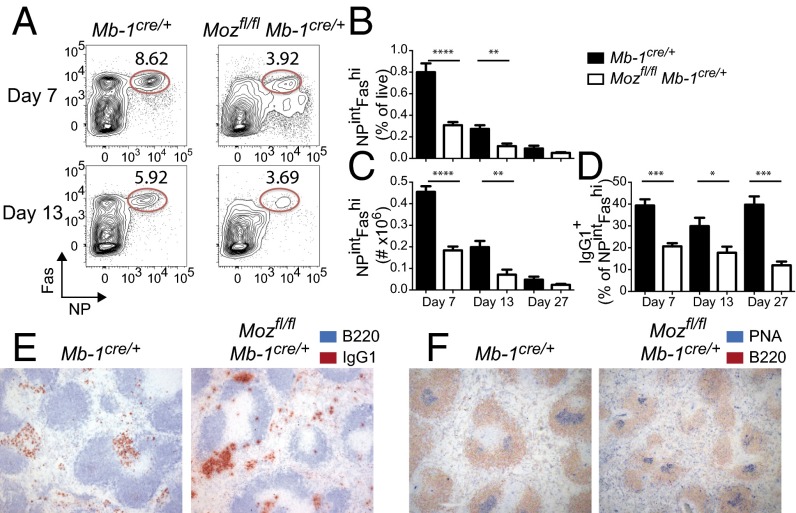
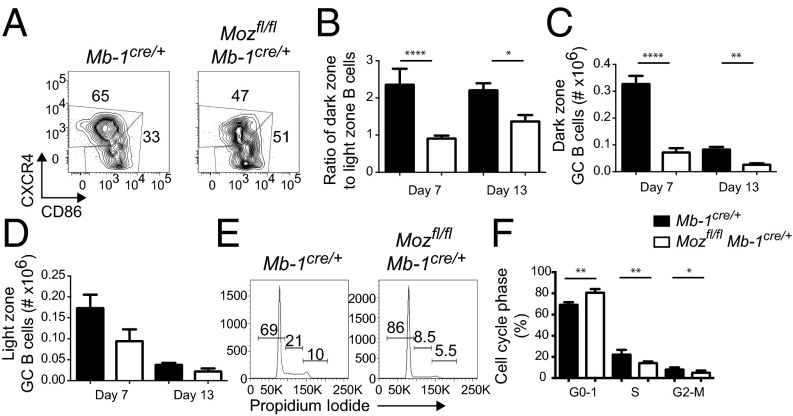
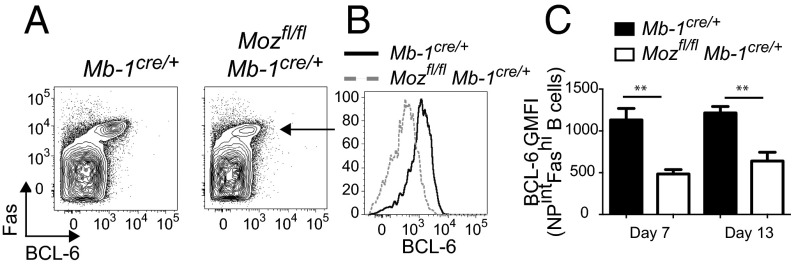
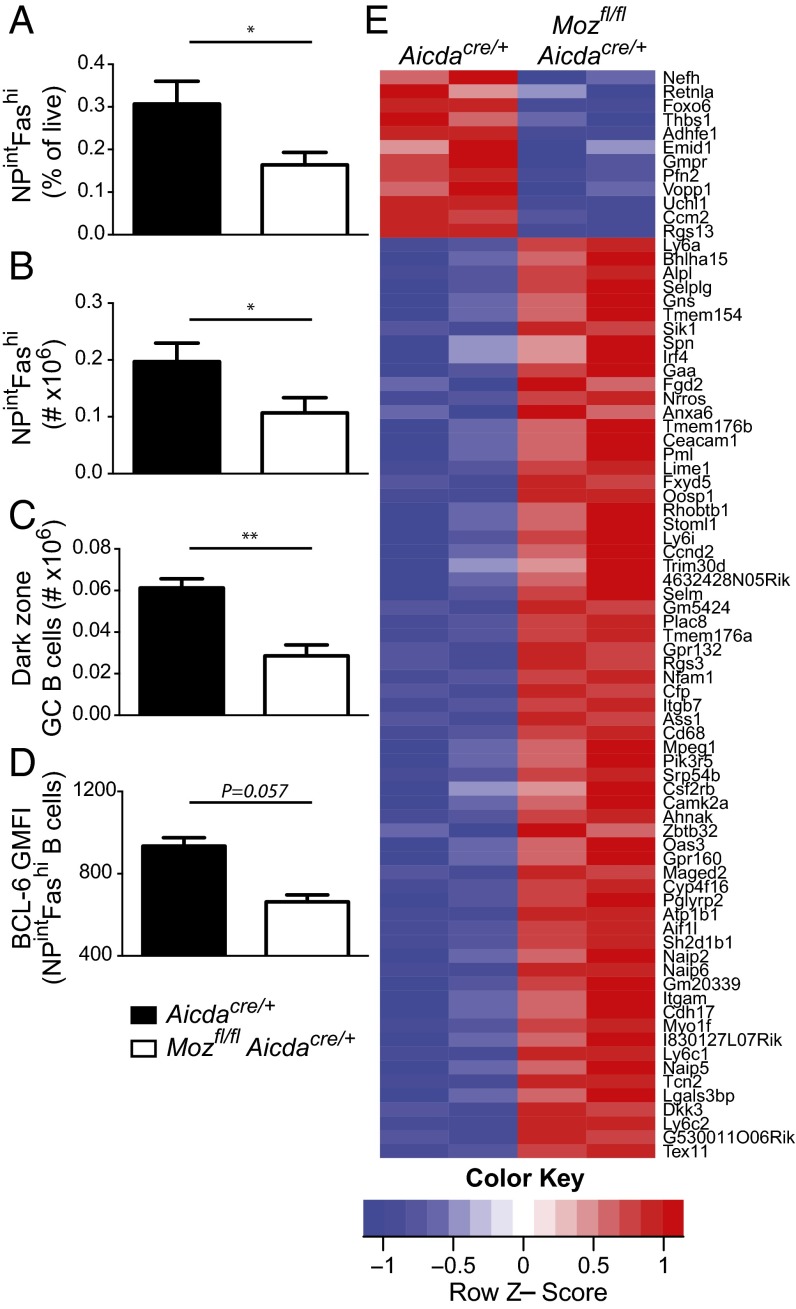
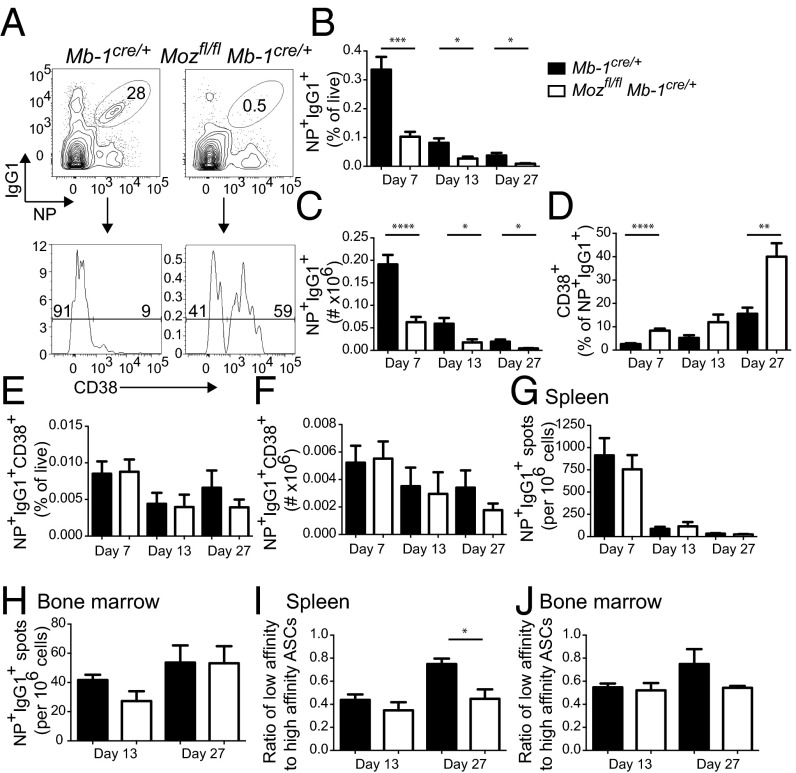
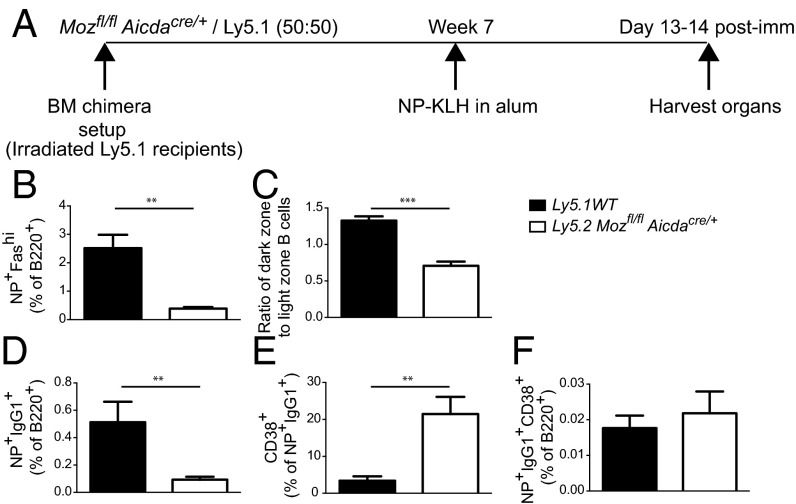
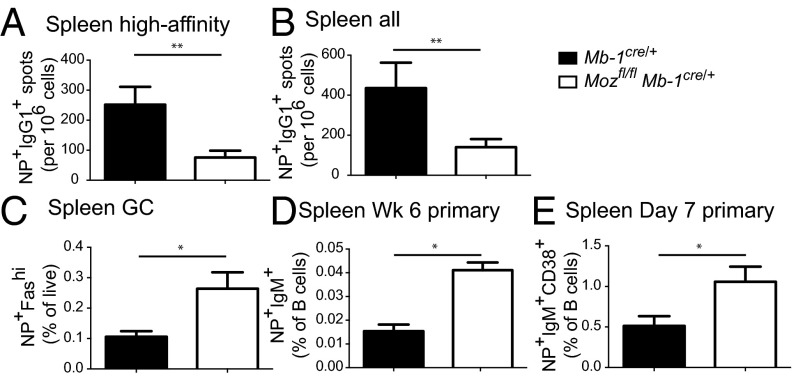
Memory B cells and long-lived bone marrow-resident plasma cells maintain humoral immunity. Little is known about the intrinsic mechanisms that are essential for forming memory B cells or endowing them with the ability to rapidly differentiate upon reexposure while maintaining the population over time. Histone modifications have been shown to regulate lymphocyte development, but their role in regulating differentiation and maintenance of B-cell subsets during an immune response is unclear. Using stage-specific deletion of monocytic leukemia zinc finger protein (MOZ), a histone acetyltransferase, we demonstrate that mutation of this chromatin modifier alters fate decisions in both primary and secondary responses. In the absence of MOZ, germinal center B cells were significantly impaired in their ability to generate dark zone centroblasts, with a concomitant decrease in both cell-cycle progression and BCL-6 expression. In contrast, there was increased differentiation to IgM and low-affinity IgG1(+) memory B cells. The lack of MOZ affected the functional outcome of humoral immune responses, with an increase in secondary germinal centers and a corresponding decrease in secondary high-affinity antibody-secreting cell formation. Therefore, these data provide strong evidence that manipulating epigenetic modifiers can regulate fate decisions during humoral responses, and thus could be targeted for therapeutic intervention.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Plexin-D1 is a novel regulator of germinal centers and humoral immune responses.J Immunol. 2011 May 15;186(10):5603-11. doi: 10.4049/jimmunol.1003464. Epub 2011 Apr 4. J Immunol. 2011. PMID: 21464091 Free PMC article.
-
B Cell-Intrinsic mTORC1 Promotes Germinal Center-Defining Transcription Factor Gene Expression, Somatic Hypermutation, and Memory B Cell Generation in Humoral Immunity.J Immunol. 2018 Apr 15;200(8):2627-2639. doi: 10.4049/jimmunol.1701321. Epub 2018 Mar 12. J Immunol. 2018. PMID: 29531165 Free PMC article.
-
High affinity IgM(+) memory B cells are generated through a germinal center-dependent pathway.Mol Immunol. 2015 Dec;68(2 Pt C):617-27. doi: 10.1016/j.molimm.2015.10.003. Epub 2015 Oct 26. Mol Immunol. 2015. PMID: 26514429
-
Plasma cell differentiation during the germinal center reaction.Immunol Rev. 2019 Mar;288(1):64-74. doi: 10.1111/imr.12751. Immunol Rev. 2019. PMID: 30874351 Review.
-
New insights into the regulation of human B-cell differentiation.Trends Immunol. 2009 Jun;30(6):277-85. doi: 10.1016/j.it.2009.03.008. Epub 2009 May 15. Trends Immunol. 2009. PMID: 19447676 Free PMC article. Review.
Cited by
-
Pre-mitotic genome re-organisation bookends the B cell differentiation process.Nat Commun. 2021 Feb 26;12(1):1344. doi: 10.1038/s41467-021-21536-2. Nat Commun. 2021. PMID: 33637722 Free PMC article.
-
Germinal Center B Cell Dynamics.Immunity. 2016 Sep 20;45(3):471-482. doi: 10.1016/j.immuni.2016.09.001. Immunity. 2016. PMID: 27653600 Free PMC article. Review.
-
The MOZ-BRPF1 acetyltransferase complex in epigenetic crosstalk linked to gene regulation, development, and human diseases.Front Cell Dev Biol. 2023 Jan 11;10:1115903. doi: 10.3389/fcell.2022.1115903. eCollection 2022. Front Cell Dev Biol. 2023. PMID: 36712963 Free PMC article. Review.
-
Epigenetics of the antibody and autoantibody response.Curr Opin Immunol. 2020 Dec;67:75-86. doi: 10.1016/j.coi.2020.09.004. Epub 2020 Nov 8. Curr Opin Immunol. 2020. PMID: 33176228 Free PMC article. Review.
-
Advances in understanding the formation and fate of B-cell memory in response to immunization or infection.Oxf Open Immunol. 2021 Sep 11;2(1):iqab018. doi: 10.1093/oxfimm/iqab018. eCollection 2021. Oxf Open Immunol. 2021. PMID: 36845573 Free PMC article. Review.
References
-
- Tarlinton D, Good-Jacobson K. Diversity among memory B cells: Origin, consequences, and utility. Science. 2013;341(6151):1205–1211. - PubMed
-
- Han S, et al. Cellular interaction in germinal centers. Roles of CD40 ligand and B7-2 in established germinal centers. J Immunol. 1995;155(2):556–567. - PubMed
-
- Jacob J, Kelsoe G, Rajewsky K, Weiss U. Intraclonal generation of antibody mutants in germinal centres. Nature. 1991;354(6352):389–392. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
