

doi: 10.1083/jcb.201401126.
Bridging the gap: membrane contact sites in signaling, metabolism, and organelle dynamics
Affiliations
- PMID: 24958771
- PMCID: PMC4068136
- DOI: 10.1083/jcb.201401126
Item in Clipboard
Bridging the gap: membrane contact sites in signaling, metabolism, and organelle dynamics
J Cell Biol.
.
Abstract
Regions of close apposition between two organelles, often referred to as membrane contact sites (MCSs), mostly form between the endoplasmic reticulum and a second organelle, although contacts between mitochondria and other organelles have also begun to be characterized. Although these contact sites have been noted since cells first began to be visualized with electron microscopy, the functions of most of these domains long remained unclear. The last few years have witnessed a dramatic increase in our understanding of MCSs, revealing the critical roles they play in intracellular signaling, metabolism, the trafficking of metabolites, and organelle inheritance, division, and transport.
Figures
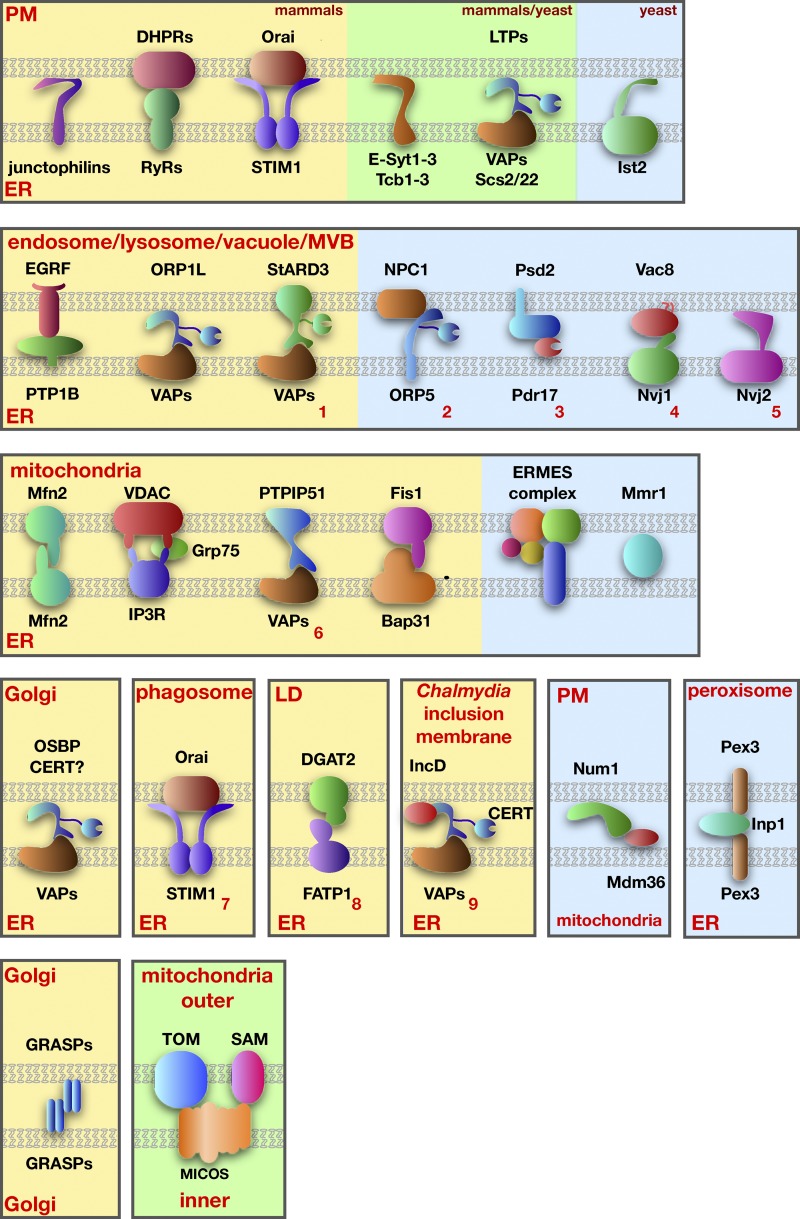
Proteins proposed to mediate tethering at MCSs. Mammalian proteins are shown on a yellow background, yeast proteins on a blue background, and proteins found in both mammals and yeast are on a green background. Tethering complexes not described in the text are indicated with red numbers: (1) StARD3-VAPs (Alpy et al., 2013), (2) NPC1-ORP5 (Du et al., 2011), (3) Psd2-Pdr17 (Riekhof et al., 2014), (4) Vac8-Nvj1 (Pan et al., 2000), (5) Nvj2 (Toulmay and Prinz, 2012), (6) PTPIP51-VAPs (De Vos et al., 2012), (7) Orai1-STIM1 (Nunes et al., 2012), (8) DGAT2-FATP1 (Xu et al., 2012), and (9) IncD-CERT-VAPs (Derré et al., 2011; Elwell et al., 2011).
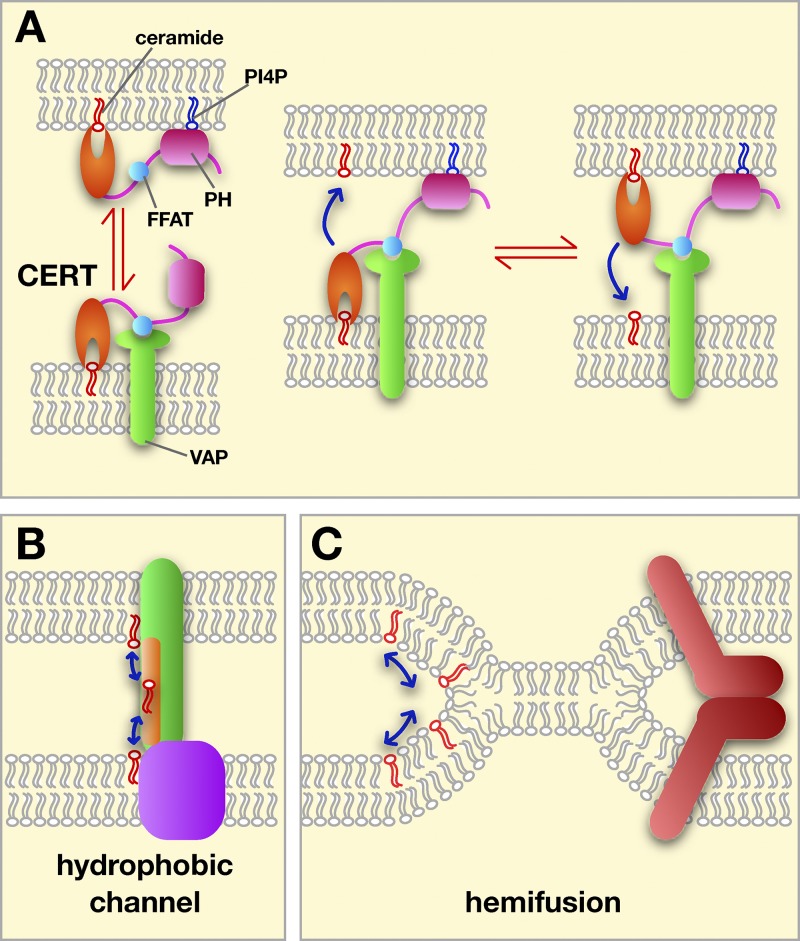
Possible mechanisms of lipid exchange at MCSs. (A) Transfer by LTPs using CERT as an example. The targeting PH domain (pink) and FFAT motif (blue) are shown. CERT could shuttle between membranes (left) or transfer while binding both membranes (right). (B) Some transfer could occur through hydrophobic channels or tunnels (in green) bridging the two membranes at a MCS. (C) Lipid exchange between hemifused membranes. Hemifusion could be promoted and regulated by proteins (red).
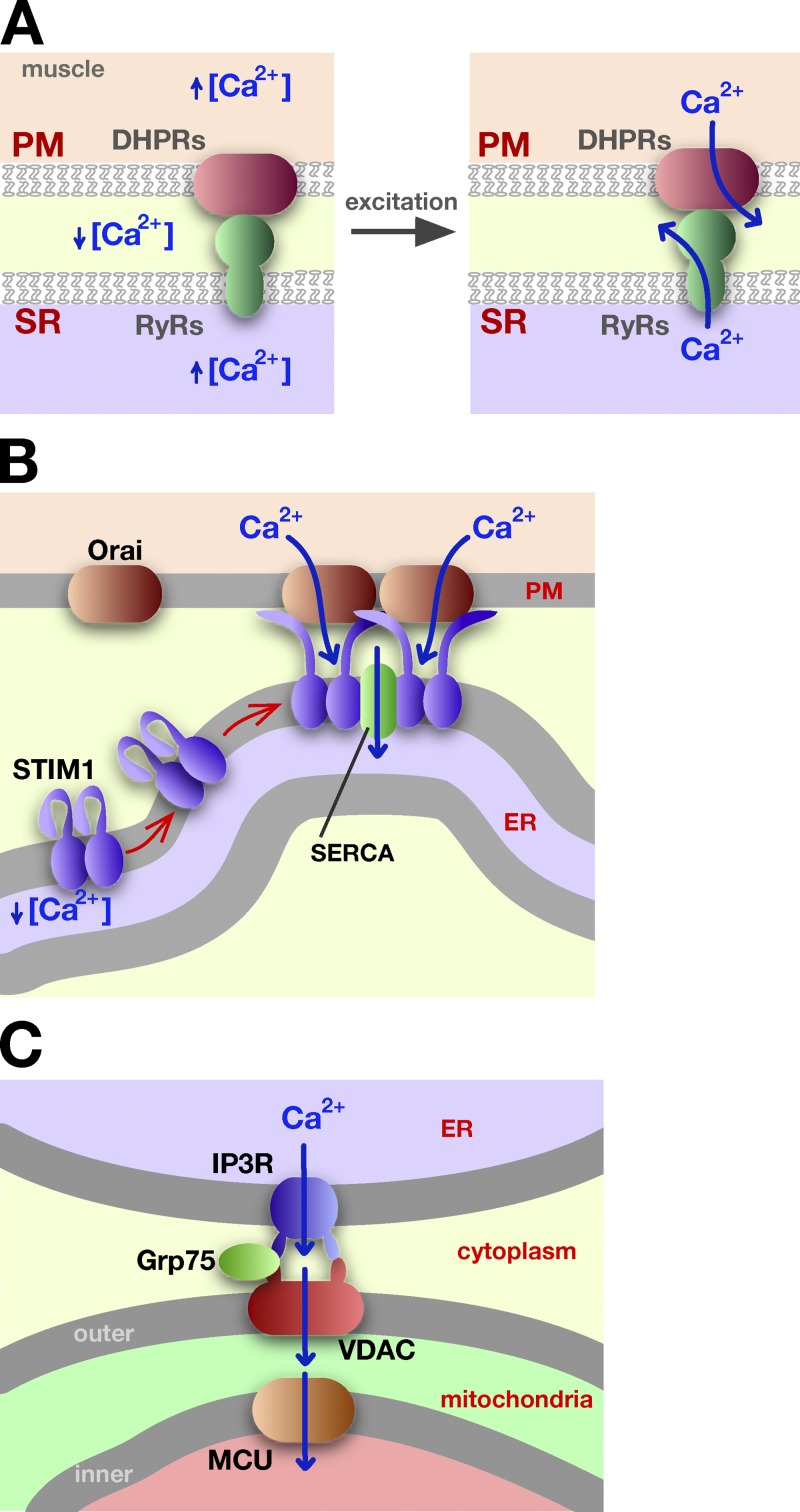
Ca2+ trafficking at ER–PM MCSs. (A) In muscle cells, the interaction of the RyR in the SR and with DHPR in the PM allows the coordinated release of Ca2+ during muscle excitation and contraction. See text for details. (B) When STIM1 senses low Ca2+ concentration in the ER, it undergoes a conformational change that allows it to oligomerize and bind to the PM, to the protein Orai1, and to accumulate at ER–PM MCSs. Ca2+ influx at these sites facilitates Ca2+ import into the ER by sarco/endoplasmic reticulum Ca2+-ATPase (SERCA). (C) Calcium channeling from the ER lumen to the mitochondrial matrix. Calcium exits the ER through the inositol trisphosphate receptor (IP3R) channel, enters mitochondria via VDAC, and then uses the mitochondrial Ca2+ uniporter (MCU) to move into the mitochondrial matrix.
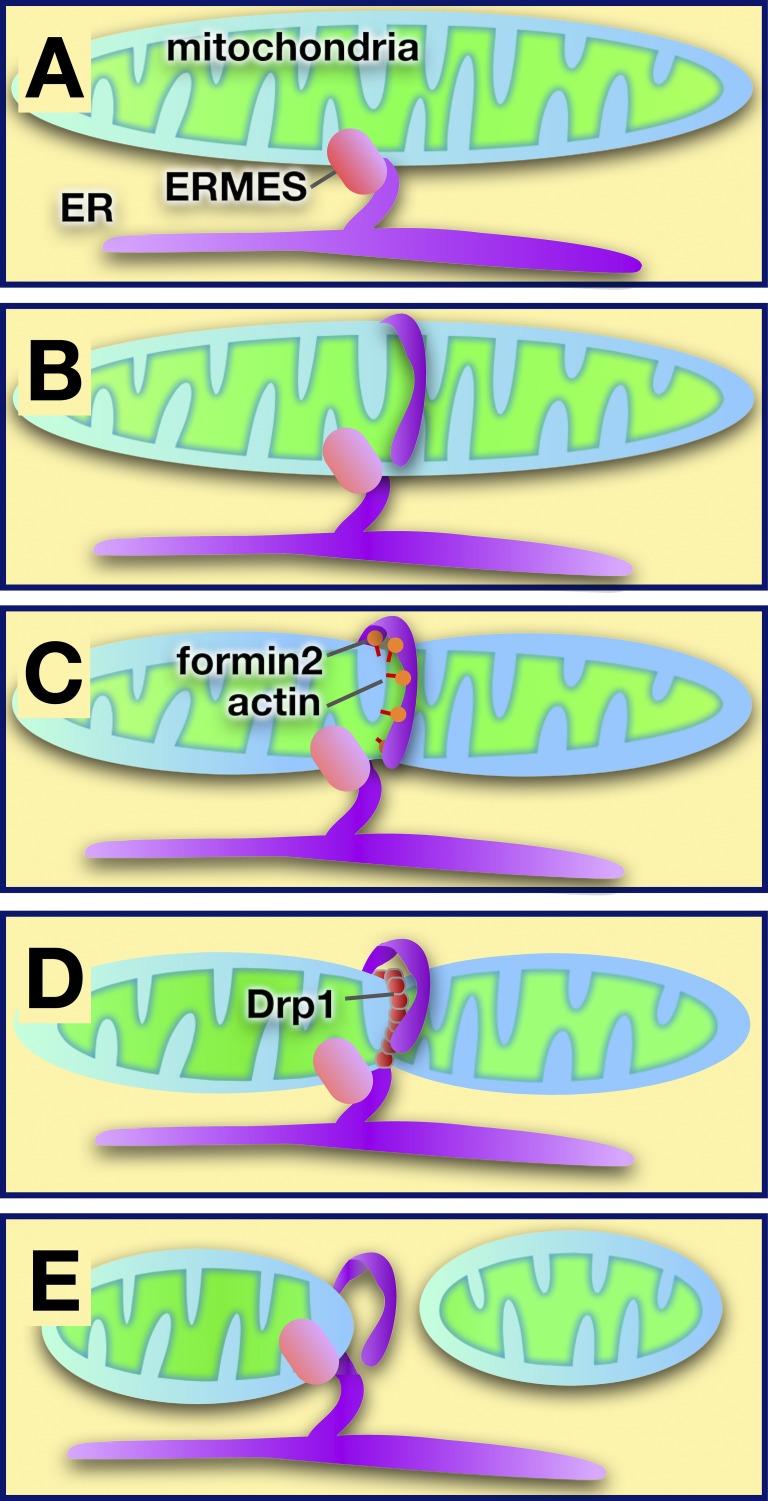
Model of ER-mediated regulation of mitochondrial fission at sites of contact. (A) The ER and mitochondria are tethered by ERMES in yeast (other tethers are used in higher eukaryotes). (B) The ER encircles mitochondria at sites where division will occur. (C) Actin polymerization facilitated by formin 2 may cause mitochondrial constriction. (D) The dynamin-like protein Drp1 is recruited to the mitochondrial surface, where it multimerizes and causes mitochondrial scission. (E) After fission, the ER remains associated with the mitochondrion that retains the ERMES complex.
Similar articles
-
Lipid transfer and signaling at organelle contact sites: the tip of the iceberg.Curr Opin Cell Biol. 2011 Aug;23(4):458-63. doi: 10.1016/j.ceb.2011.04.006. Curr Opin Cell Biol. 2011. PMID: 21555211 Free PMC article. Review.
-
Interacting organelles.Curr Opin Cell Biol. 2018 Aug;53:84-91. doi: 10.1016/j.ceb.2018.06.003. Epub 2018 Jul 2. Curr Opin Cell Biol. 2018. PMID: 30006038 Free PMC article. Review.
-
Organelle remodeling at membrane contact sites.J Struct Biol. 2016 Oct;196(1):15-19. doi: 10.1016/j.jsb.2016.05.003. Epub 2016 May 13. J Struct Biol. 2016. PMID: 27181417 Free PMC article. Review.
-
Short-range intracellular trafficking of small molecules across endoplasmic reticulum junctions.Trends Cell Biol. 2004 Sep;14(9):483-90. doi: 10.1016/j.tcb.2004.07.017. Trends Cell Biol. 2004. PMID: 15350976
-
Membrane contact sites, gateways for lipid homeostasis.Curr Opin Cell Biol. 2015 Apr;33:82-87. doi: 10.1016/j.ceb.2014.12.004. Epub 2015 Jan 6. Curr Opin Cell Biol. 2015. PMID: 25569848 Free PMC article. Review.
Cited by
-
Uptake and Metabolic Conversion of Exogenous Phosphatidylcholines Depending on Their Acyl Chain Structure in Arabidopsis thaliana.Int J Mol Sci. 2023 Dec 20;25(1):89. doi: 10.3390/ijms25010089. Int J Mol Sci. 2023. PMID: 38203257 Free PMC article.
-
Lipid Metabolism in Macrophages: Focus on Atherosclerosis.Biomedicines. 2020 Aug 1;8(8):262. doi: 10.3390/biomedicines8080262. Biomedicines. 2020. PMID: 32752275 Free PMC article. Review.
-
Dynamics and functions of lipid droplets.Nat Rev Mol Cell Biol. 2019 Mar;20(3):137-155. doi: 10.1038/s41580-018-0085-z. Nat Rev Mol Cell Biol. 2019. PMID: 30523332 Free PMC article. Review.
-
Ca2+ Dyshomeostasis Links Risk Factors to Neurodegeneration in Parkinson's Disease.Front Cell Neurosci. 2022 Apr 14;16:867385. doi: 10.3389/fncel.2022.867385. eCollection 2022. Front Cell Neurosci. 2022. PMID: 35496903 Free PMC article. Review.
-
The Expanding and Unexpected Functions of Mitochondria Contact Sites.Trends Cell Biol. 2019 Jul;29(7):580-590. doi: 10.1016/j.tcb.2019.02.009. Epub 2019 Mar 28. Trends Cell Biol. 2019. PMID: 30929794 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
