

Identification of fatty acid binding protein 4 as an adipokine that regulates insulin secretion during obesity
- PMID: 24944906
- PMCID: PMC4060222
- DOI: 10.1016/j.molmet.2014.02.005
Identification of fatty acid binding protein 4 as an adipokine that regulates insulin secretion during obesity
Abstract
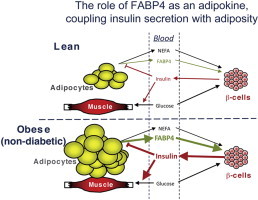
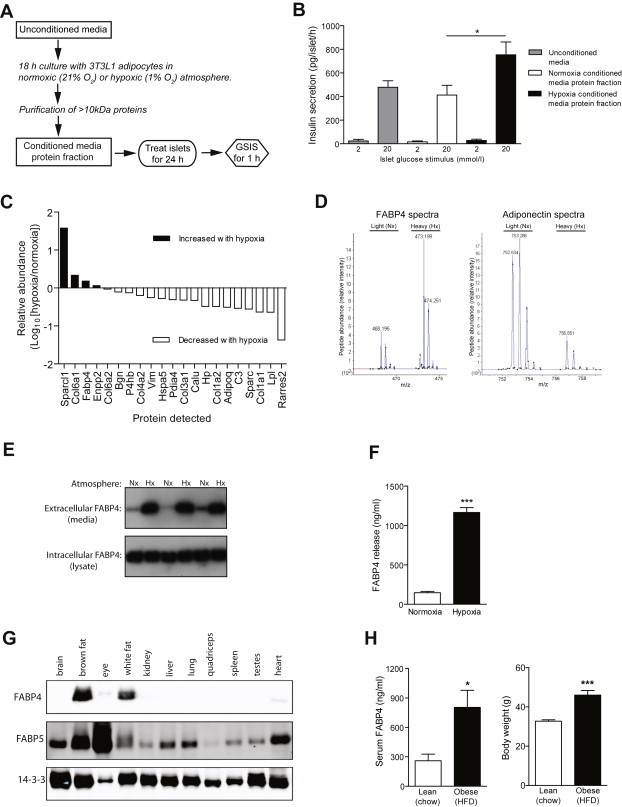
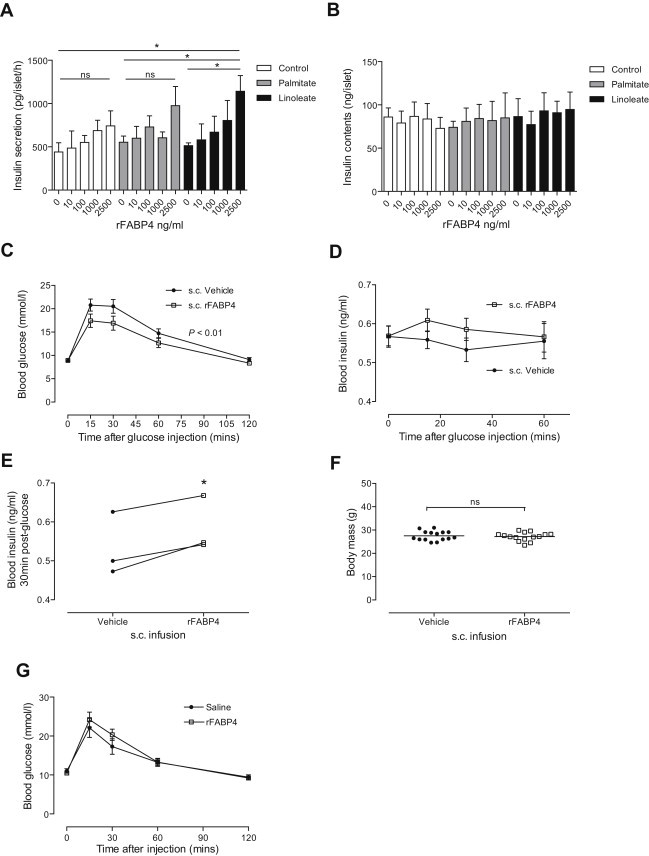
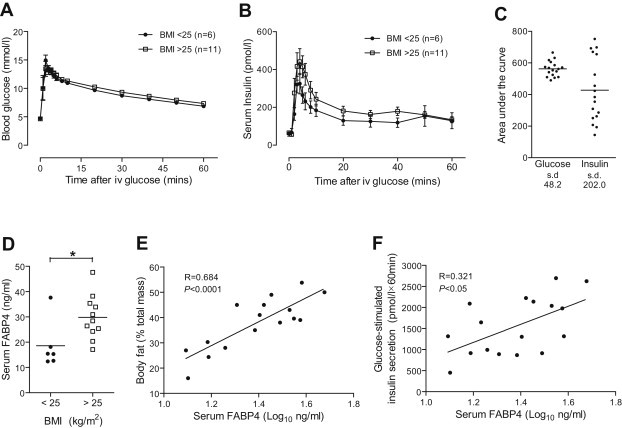
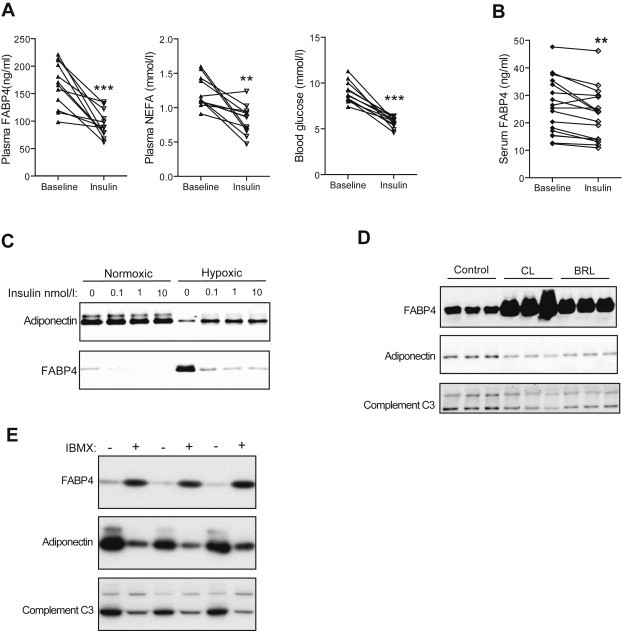
A critical feature of obesity is enhanced insulin secretion from pancreatic β-cells, enabling the majority of individuals to maintain glycaemic control despite adiposity and insulin resistance. Surprisingly, the factors coordinating this adaptive β-cell response with adiposity have not been delineated. Here we show that fatty acid binding protein 4 (FABP4/aP2) is an adipokine released from adipocytes under obesogenic conditions, such as hypoxia, to augment insulin secretion. The insulinotropic action of FABP4 was identified using an in vitro system that recapitulates adipocyte to β-cell endocrine signalling, with glucose-stimulated insulin secretion (GSIS) as a functional readout, coupled with quantitative proteomics. Exogenous FABP4 potentiated GSIS in vitro and in vivo, and circulating FABP4 levels correlated with GSIS in humans. Insulin inhibited FABP4 release from adipocytes in vitro, in mice and in humans, consistent with feedback regulation. These data suggest that FABP4 and insulin form an endocrine loop coordinating the β-cell response to obesity.
Keywords: Adipocyte; Adipokine; BMI, body mass index; Beta-cell; ELISA, enzyme-linked immunosorbant assay; FABP4; GSIS, glucose-stimulated insulin secretion; IBMX, 3-Isobutyl-1-methylxanthine; Insulin secretion; NEFA, non-esterified fatty acid; Obesity; SILAC, stable-isotope labelling by amino acids in cell culture; T2D, type 2 diabetes; cAMP, cyclic-AMP.
Figures
Similar articles
-
Angiotensin II receptor blockers decrease serum concentration of fatty acid-binding protein 4 in patients with hypertension.Hypertens Res. 2015 Apr;38(4):252-9. doi: 10.1038/hr.2015.2. Epub 2015 Feb 12. Hypertens Res. 2015. PMID: 25672659
-
The control of insulin secretion by adipokines: current evidence for adipocyte-beta cell endocrine signalling in metabolic homeostasis.Mamm Genome. 2014 Oct;25(9-10):442-54. doi: 10.1007/s00335-014-9538-7. Epub 2014 Aug 22. Mamm Genome. 2014. PMID: 25146550 Review.
-
Beta cell compensation for insulin resistance in Zucker fatty rats: increased lipolysis and fatty acid signalling.Diabetologia. 2006 Sep;49(9):2120-30. doi: 10.1007/s00125-006-0305-5. Epub 2006 Jul 26. Diabetologia. 2006. PMID: 16868750
-
Reduction of serum FABP4 level by sitagliptin, a DPP-4 inhibitor, in patients with type 2 diabetes mellitus.J Lipid Res. 2015 Dec;56(12):2372-80. doi: 10.1194/jlr.M059469. Epub 2015 Oct 14. J Lipid Res. 2015. PMID: 26467280 Free PMC article.
-
Associations between Fatty Acid-Binding Protein 4⁻A Proinflammatory Adipokine and Insulin Resistance, Gestational and Type 2 Diabetes Mellitus.Cells. 2019 Mar 8;8(3):227. doi: 10.3390/cells8030227. Cells. 2019. PMID: 30857223 Free PMC article. Review.
Cited by
-
Unconventional Secretion of Adipocyte Fatty Acid Binding Protein 4 Is Mediated By Autophagic Proteins in a Sirtuin-1-Dependent Manner.Diabetes. 2019 Sep;68(9):1767-1777. doi: 10.2337/db18-1367. Epub 2019 Jun 6. Diabetes. 2019. PMID: 31171562 Free PMC article.
-
Beta Cell Function and the Nutritional State: Dietary Factors that Influence Insulin Secretion.Curr Diab Rep. 2015 Oct;15(10):76. doi: 10.1007/s11892-015-0650-1. Curr Diab Rep. 2015. PMID: 26294335 Review.
-
A-FABP mediates adaptive thermogenesis by promoting intracellular activation of thyroid hormones in brown adipocytes.Nat Commun. 2017 Jan 27;8:14147. doi: 10.1038/ncomms14147. Nat Commun. 2017. PMID: 28128199 Free PMC article.
-
Adipose Signals Regulating Distal Organ Health and Disease.Diabetes. 2024 Feb 1;73(2):169-177. doi: 10.2337/dbi23-0005. Diabetes. 2024. PMID: 38241508 Free PMC article. Review.
-
FABP4 contributes to renal interstitial fibrosis via mediating inflammation and lipid metabolism.Cell Death Dis. 2019 May 16;10(6):382. doi: 10.1038/s41419-019-1610-5. Cell Death Dis. 2019. Retraction in: Cell Death Dis. 2021 Aug 20;12(9):801. doi: 10.1038/s41419-021-04084-x. PMID: 31097687 Free PMC article. Retracted.
References
-
- Yach D., Stuckler D., Brownell K.D. Epidemiologic and economic consequences of the global epidemics of obesity and diabetes. Nature Medicine. 2006;12:62–66. - PubMed
-
- Zimmet P., Alberti K.G., Shaw J. Global and societal implications of the diabetes epidemic. Nature. 2001;414:782–787. - PubMed
-
- Eckel R.H., Grundy S.M., Zimmet P.Z. The metabolic syndrome. Lancet. 2005;365:1415–1428. - PubMed
-
- Alberti K.G.M.M., Eckel R.H., Grundy S.M., Zimmet P.Z., Cleeman J.I., Donato K.A. Harmonizing the metabolic syndrome. Circulation. 2009;120:1640–1645. - PubMed
-
- Calle E.E., Thun M.J. Obesity and cancer. Oncogene. 2004;23:6365–6378. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
