

Transcriptional regulation by Pho23 modulates the frequency of autophagosome formation
- PMID: 24881874
- PMCID: PMC4169046
- DOI: 10.1016/j.cub.2014.04.048
Transcriptional regulation by Pho23 modulates the frequency of autophagosome formation
Abstract
Background: Autophagy as a conserved lysosomal/vacuolar degradation and recycling pathway is important in normal development and physiology, and defects in this process are linked to many kinds of disease. Because too much or too little autophagy can be detrimental, the process must be tightly regulated both temporally and in magnitude. Two parameters that affect this regulation are the size and the number of autophagosomes; however, although we know that the amount of Atg8 affects the size of autophagosomes, the mechanism for regulating their number has not been elucidated. The transcriptional induction and repression of the autophagy-related (ATG) genes is one crucial aspect of autophagy regulation, but the transcriptional regulators that modulate autophagy are not well characterized.
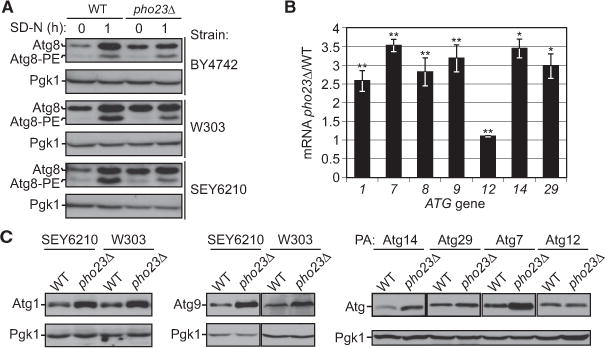
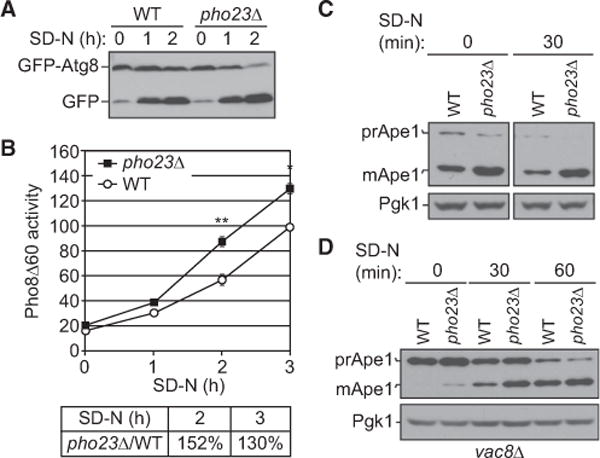
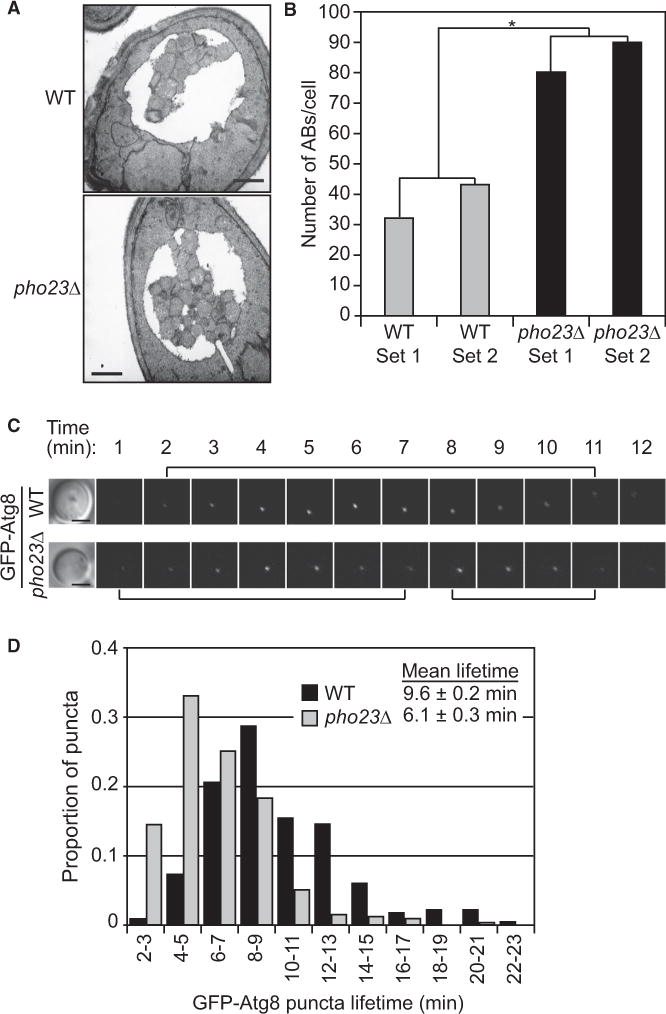
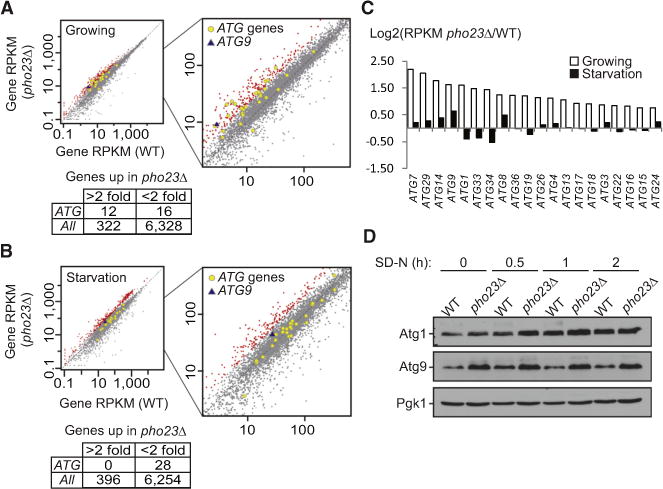
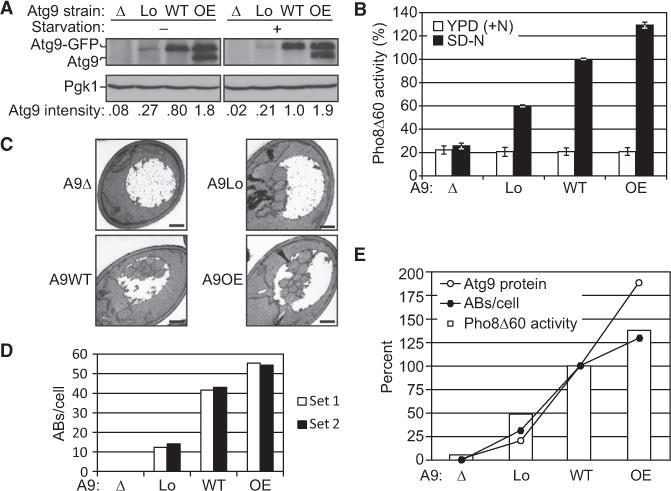
Results: We detected increased expression levels of ATG genes, and elevated autophagy activity, in cells lacking the transcriptional regulator Pho23. Using transmission electron microscopy, we found that PHO23 null mutant cells contain significantly more autophagosomes than the wild-type. By RNA sequencing transcriptome profiling, we identified ATG9 as one of the key targets of Pho23, and our studies with strains expressing modulated levels of Atg9 show that the amount of this protein directly correlates with the frequency of autophagosome formation and the level of autophagy activity.
Conclusions: Our results identified Pho23 as a master transcriptional repressor for autophagy that regulates the frequency of autophagosome formation through its negative regulation of ATG9.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Transcriptional regulation of ATG9 by the Pho23-Rpd3 complex modulates the frequency of autophagosome formation.Autophagy. 2014 Sep;10(9):1681-2. doi: 10.4161/auto.29641. Epub 2014 Jul 7. Autophagy. 2014. PMID: 25046109 Free PMC article. Review.
-
A nuclear membrane-derived structure associated with Atg8 is involved in the sequestration of selective cargo, the Cvt complex, during autophagosome formation in yeast.Autophagy. 2019 Mar;15(3):423-437. doi: 10.1080/15548627.2018.1525475. Epub 2018 Oct 11. Autophagy. 2019. PMID: 30238844 Free PMC article.
-
Atg9-centered multi-omics integration reveals new autophagy regulators in Saccharomyces cerevisiae.Autophagy. 2021 Dec;17(12):4453-4476. doi: 10.1080/15548627.2021.1898749. Epub 2021 Mar 15. Autophagy. 2021. PMID: 33722159 Free PMC article.
-
Atg27 is required for autophagy-dependent cycling of Atg9.Mol Biol Cell. 2007 Feb;18(2):581-93. doi: 10.1091/mbc.e06-07-0612. Epub 2006 Nov 29. Mol Biol Cell. 2007. PMID: 17135291 Free PMC article.
-
Atg9 trafficking in the yeast Saccharomyces cerevisiae.Autophagy. 2007 Mar-Apr;3(2):145-8. doi: 10.4161/auto.3608. Epub 2007 Mar 21. Autophagy. 2007. PMID: 17204846 Review.
Cited by
-
A COPII subunit acts with an autophagy receptor to target endoplasmic reticulum for degradation.Science. 2019 Jul 5;365(6448):53-60. doi: 10.1126/science.aau9263. Science. 2019. PMID: 31273116 Free PMC article.
-
Identification of transcription factors that regulate ATG8 expression and autophagy in Arabidopsis.Autophagy. 2020 Jan;16(1):123-139. doi: 10.1080/15548627.2019.1598753. Epub 2019 Apr 6. Autophagy. 2020. PMID: 30909785 Free PMC article.
-
Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition).Autophagy. 2016;12(1):1-222. doi: 10.1080/15548627.2015.1100356. Autophagy. 2016. PMID: 26799652 Free PMC article. No abstract available.
-
Mechanisms of Autophagy Initiation.Annu Rev Biochem. 2017 Jun 20;86:225-244. doi: 10.1146/annurev-biochem-061516-044820. Epub 2017 Mar 15. Annu Rev Biochem. 2017. PMID: 28301741 Free PMC article. Review.
-
Ubiquitin-proteasome-mediated cyclin C degradation promotes cell survival following nitrogen starvation.Mol Biol Cell. 2020 May 1;31(10):1015-1031. doi: 10.1091/mbc.E19-11-0622. Epub 2020 Mar 11. Mol Biol Cell. 2020. PMID: 32160104 Free PMC article.
References
-
- Tsukada M, Ohsumi Y. Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Lett. 1993;333:169–174. - PubMed
-
- Huang J, Klionsky DJ. Autophagy and human disease. Cell Cycle. 2007;6:1837–1849. - PubMed
-
- Xie Z, Klionsky DJ. Autophagosome formation: core machinery and adaptations. Nat Cell Biol. 2007;9:1102–1109. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
