

A monoclonal antibody that targets a NaV1.7 channel voltage sensor for pain and itch relief
- PMID: 24856969
- PMCID: PMC4098795
- DOI: 10.1016/j.cell.2014.03.064
A monoclonal antibody that targets a NaV1.7 channel voltage sensor for pain and itch relief
Retraction in
-
Retraction Notice to: A Monoclonal Antibody that Targets a NaV1.7 Channel Voltage Sensor for Pain and Itch Relief.Cell. 2020 Jun 25;181(7):1695. doi: 10.1016/j.cell.2020.06.019. Cell. 2020. PMID: 32589960 Free PMC article. No abstract available.
Abstract
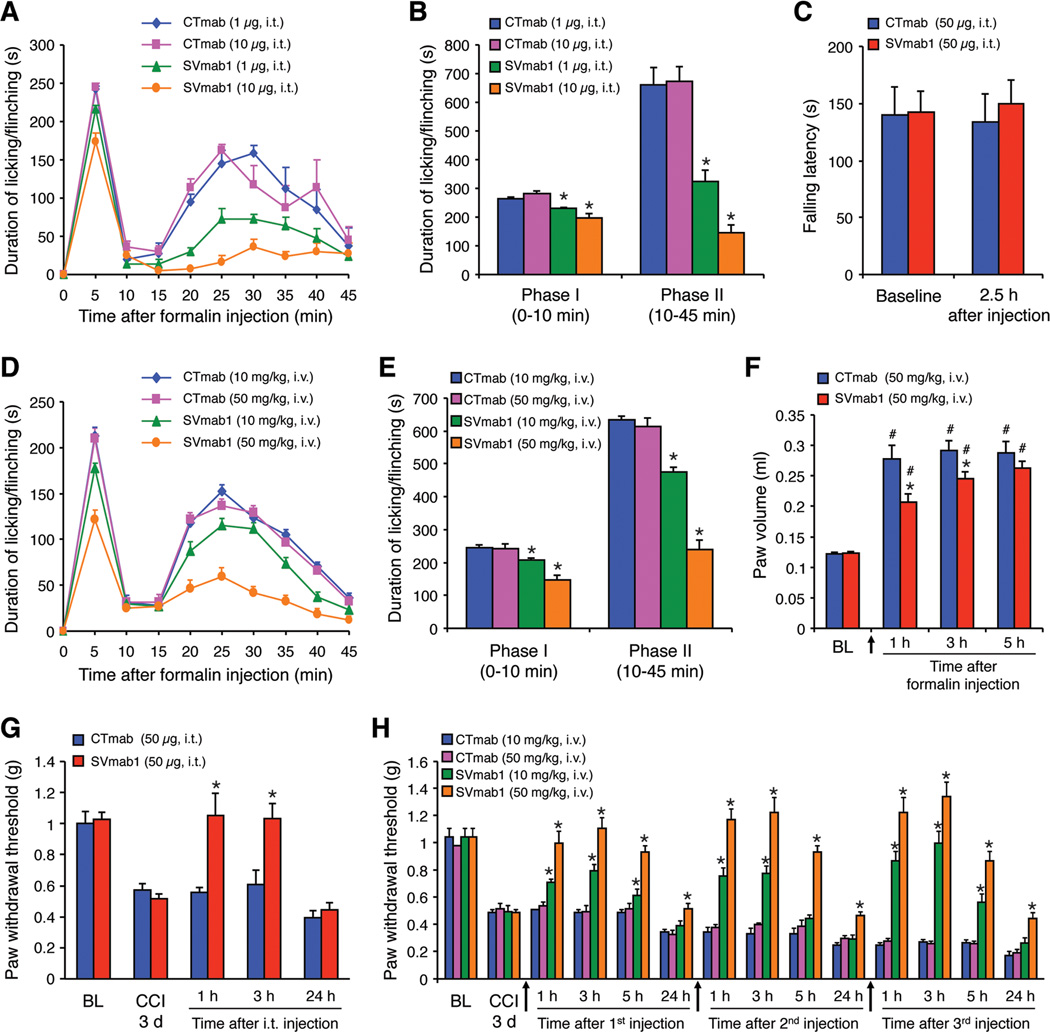
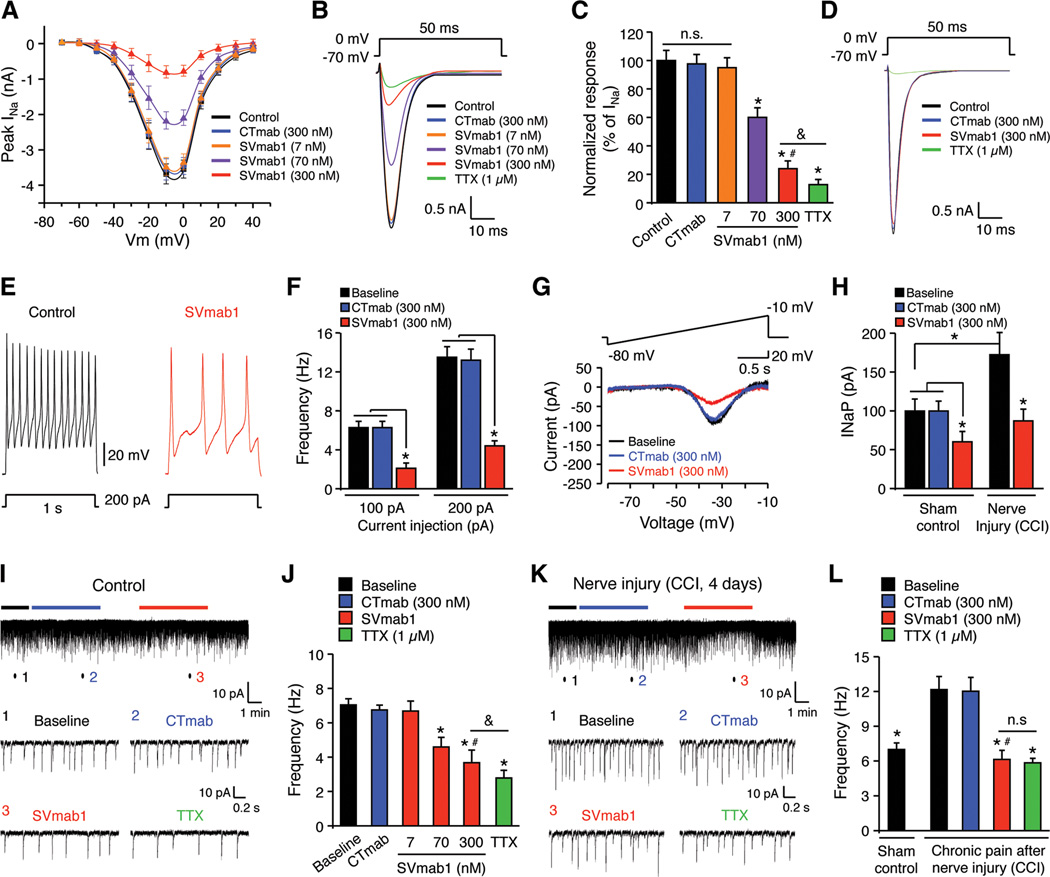
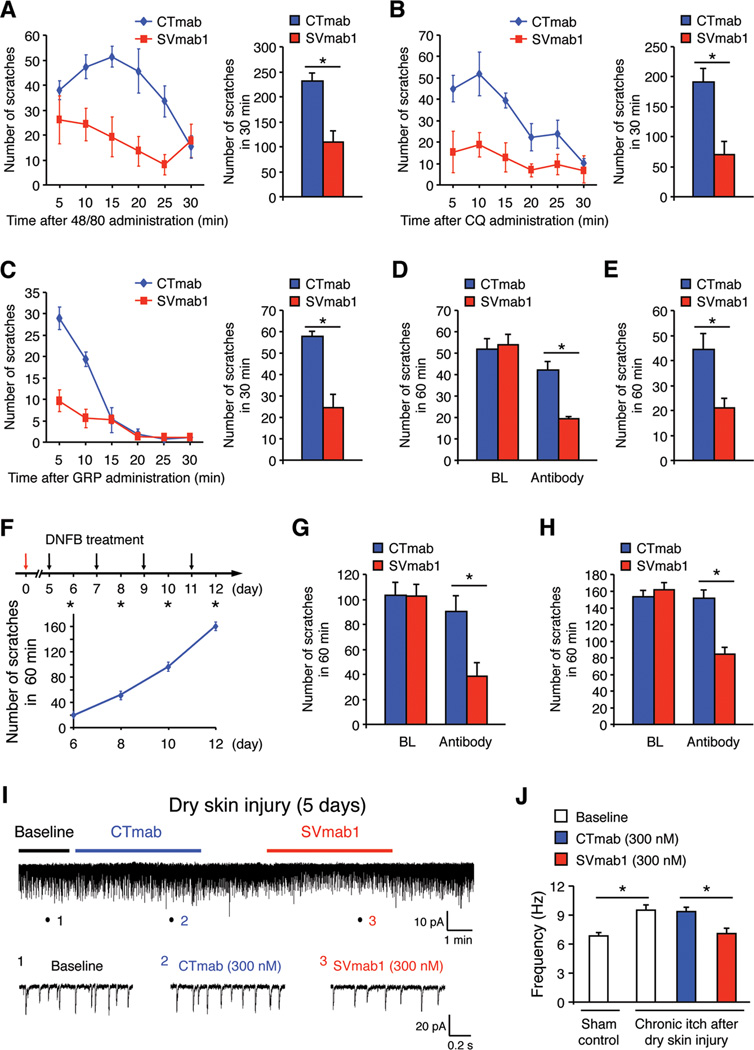
Voltage-gated sodium (NaV) channels control the upstroke of the action potentials in excitable cells. Multiple studies have shown distinct roles of NaV channel subtypes in human physiology and diseases, but subtype-specific therapeutics are lacking and the current efforts have been limited to small molecules. Here, we present a monoclonal antibody that targets the voltage-sensor paddle of NaV1.7, the subtype critical for pain sensation. This antibody not only inhibits NaV1.7 with high selectivity, but also effectively suppresses inflammatory and neuropathic pain in mice. Interestingly, the antibody inhibits acute and chronic itch despite well-documented differences in pain and itch modulation. Using this antibody, we discovered that NaV1.7 plays a key role in spinal cord nociceptive and pruriceptive synaptic transmission. Our studies reveal that NaV1.7 is a target for itch management, and the antibody has therapeutic potential for suppressing pain and itch. Our antibody strategy may have broad applications for voltage-gated cation channels.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
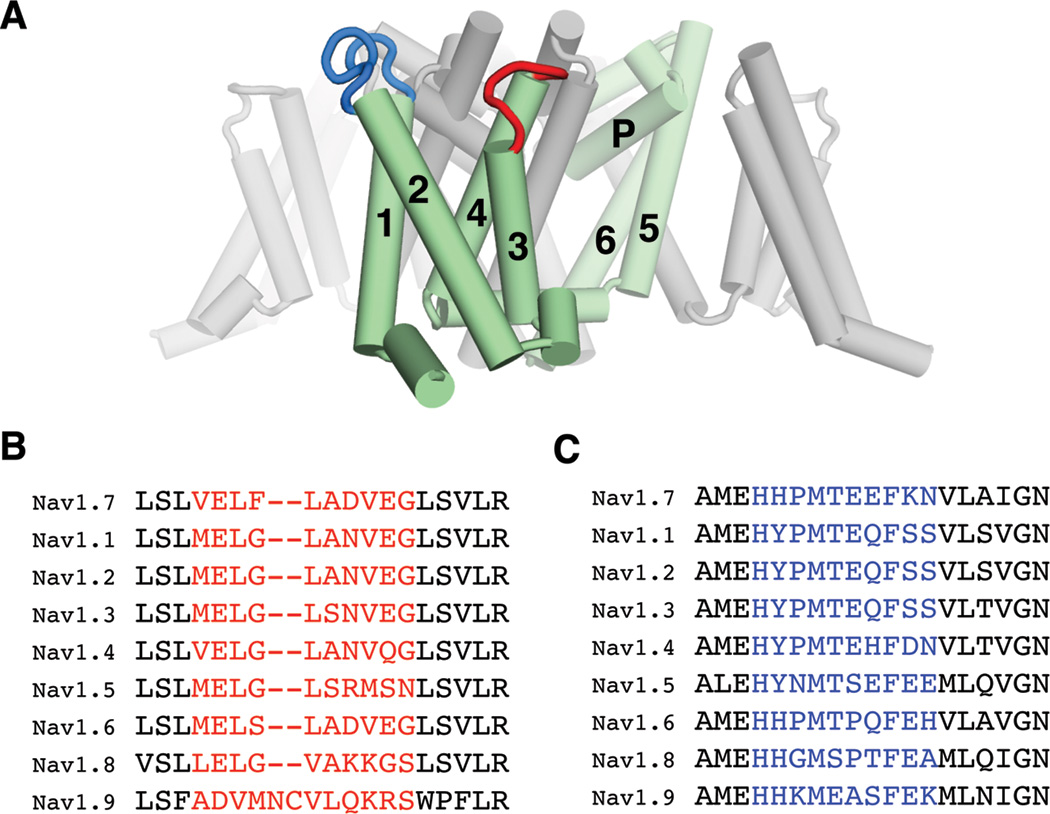
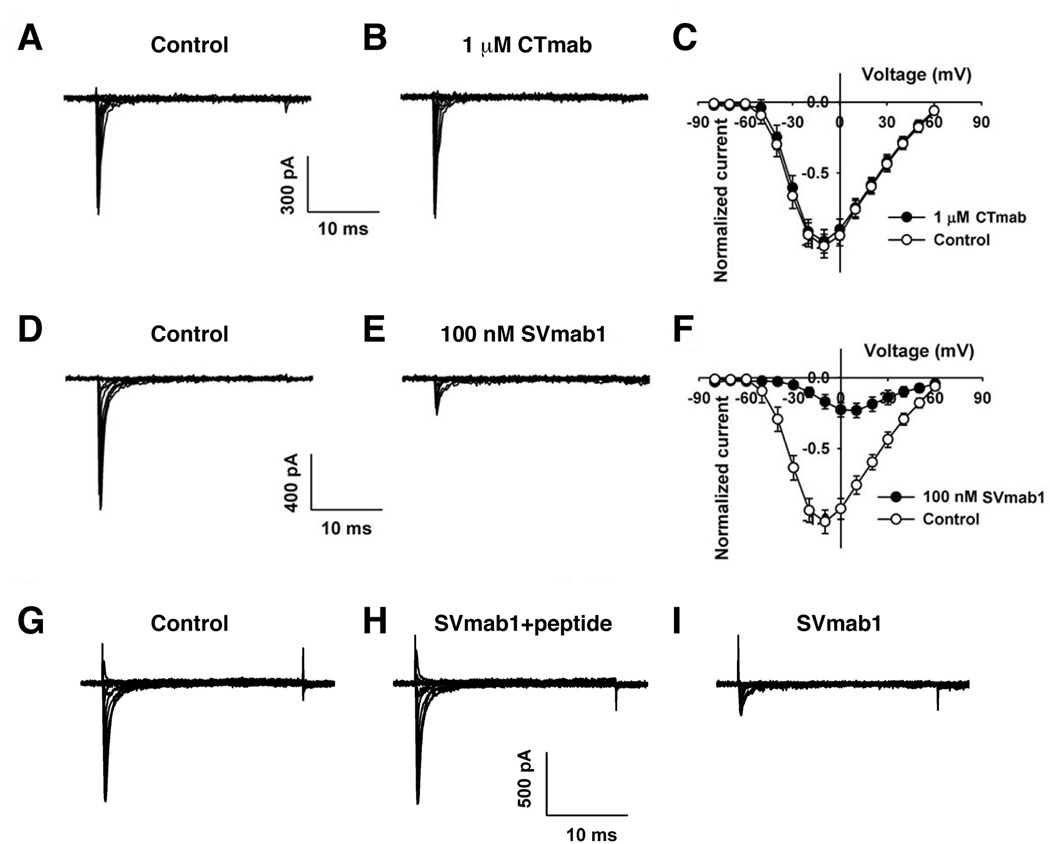
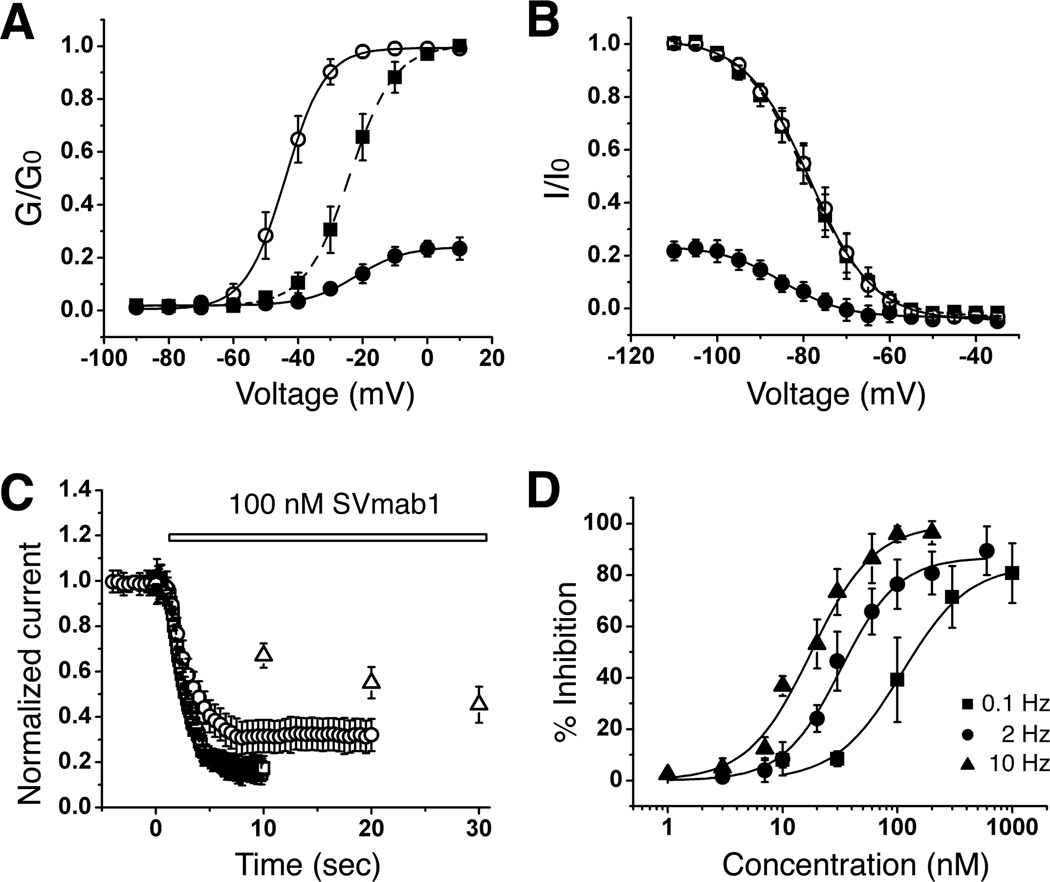
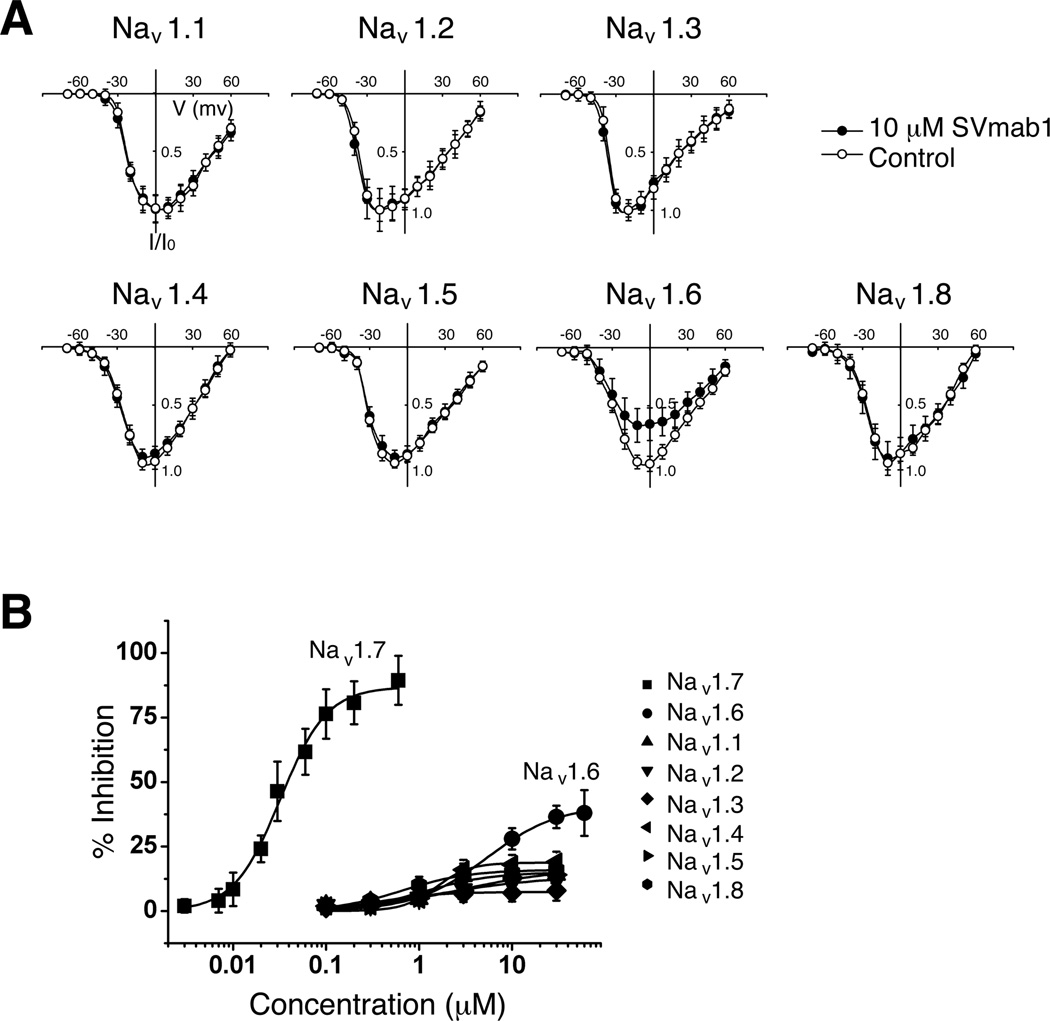
Comment in
-
Analgesia: anti-itch and anti-ouch antibody.Nat Rev Drug Discov. 2014 Aug;13(8):574. doi: 10.1038/nrd4396. Epub 2014 Jul 18. Nat Rev Drug Discov. 2014. PMID: 25033738 No abstract available.
Similar articles
-
Analgesia: anti-itch and anti-ouch antibody.Nat Rev Drug Discov. 2014 Aug;13(8):574. doi: 10.1038/nrd4396. Epub 2014 Jul 18. Nat Rev Drug Discov. 2014. PMID: 25033738 No abstract available.
-
3'-O-Methylorobol Inhibits the Voltage-Gated Sodium Channel Nav1.7 with Anti-Itch Efficacy in A Histamine-Dependent Itch Mouse Model.Int J Mol Sci. 2019 Dec 1;20(23):6058. doi: 10.3390/ijms20236058. Int J Mol Sci. 2019. PMID: 31805638 Free PMC article.
-
Inhibition of Nav1.7 channel by a novel blocker QLS-81 for alleviation of neuropathic pain.Acta Pharmacol Sin. 2021 Aug;42(8):1235-1247. doi: 10.1038/s41401-021-00682-9. Epub 2021 Jun 8. Acta Pharmacol Sin. 2021. PMID: 34103689 Free PMC article.
-
Recent advances in small molecule Nav 1.7 inhibitors for cancer pain management.Bioorg Chem. 2024 Sep;150:107605. doi: 10.1016/j.bioorg.2024.107605. Epub 2024 Jun 29. Bioorg Chem. 2024. PMID: 38971095 Review.
-
Selective Ligands and Drug Discovery Targeting the Voltage-Gated Sodium Channel Nav1.7.Handb Exp Pharmacol. 2018;246:271-306. doi: 10.1007/164_2018_97. Handb Exp Pharmacol. 2018. PMID: 29532179 Review.
Cited by
-
Transient Receptor Potential Vanilloid 4 Ion Channel Functions as a Pruriceptor in Epidermal Keratinocytes to Evoke Histaminergic Itch.J Biol Chem. 2016 May 6;291(19):10252-62. doi: 10.1074/jbc.M116.716464. Epub 2016 Mar 9. J Biol Chem. 2016. PMID: 26961876 Free PMC article.
-
Affimer proteins are versatile and renewable affinity reagents.Elife. 2017 Jun 27;6:e24903. doi: 10.7554/eLife.24903. Elife. 2017. PMID: 28654419 Free PMC article.
-
Engineering Highly Potent and Selective Microproteins against Nav1.7 Sodium Channel for Treatment of Pain.J Biol Chem. 2016 Jul 1;291(27):13974-13986. doi: 10.1074/jbc.M116.725978. Epub 2016 Apr 22. J Biol Chem. 2016. PMID: 27129258 Free PMC article.
-
Anti-nerve growth factor in pain management: current evidence.J Pain Res. 2016 Jun 8;9:373-83. doi: 10.2147/JPR.S89061. eCollection 2016. J Pain Res. 2016. PMID: 27354823 Free PMC article. Review.
-
Neurological Disturbances of Ciguatera Poisoning: Clinical Features and Pathophysiological Basis.Cells. 2020 Oct 14;9(10):2291. doi: 10.3390/cells9102291. Cells. 2020. PMID: 33066435 Free PMC article. Review.
References
-
- Beck A, Wurch T, Bailly C, Corvaia N. Strategies and challenges for the next generation of therapeutic antibodies. Nature reviews Immunology. 2010;10:345–352. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
