

Fungal chitin dampens inflammation through IL-10 induction mediated by NOD2 and TLR9 activation
- PMID: 24722226
- PMCID: PMC3983064
- DOI: 10.1371/journal.ppat.1004050
Fungal chitin dampens inflammation through IL-10 induction mediated by NOD2 and TLR9 activation
Abstract
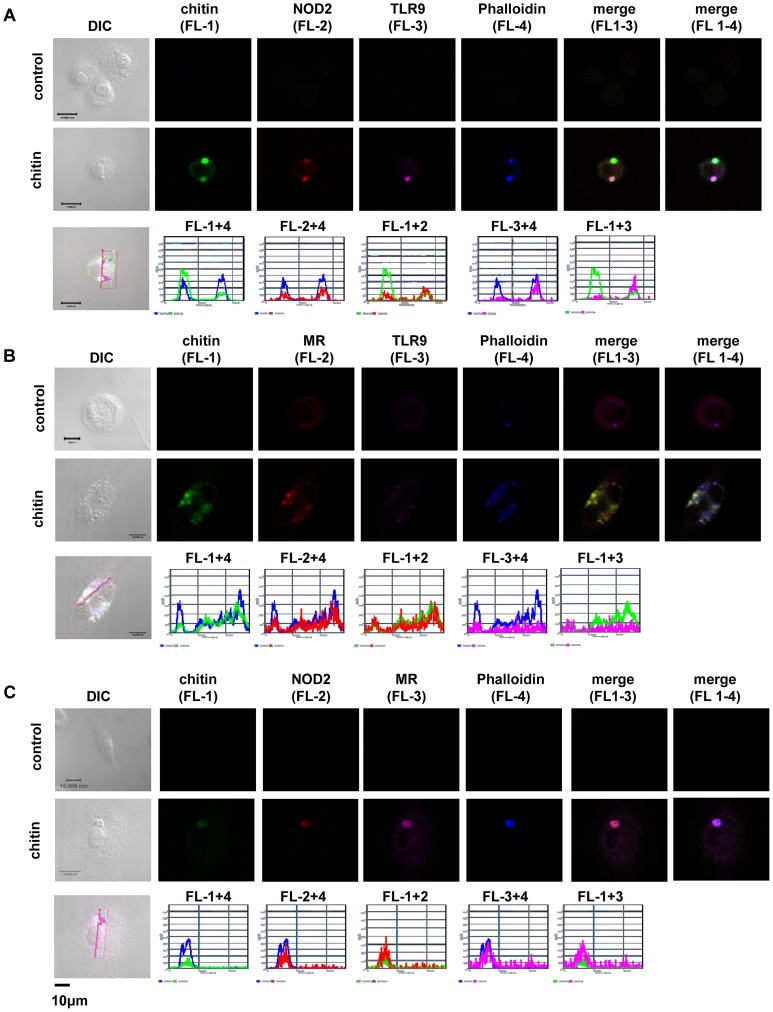
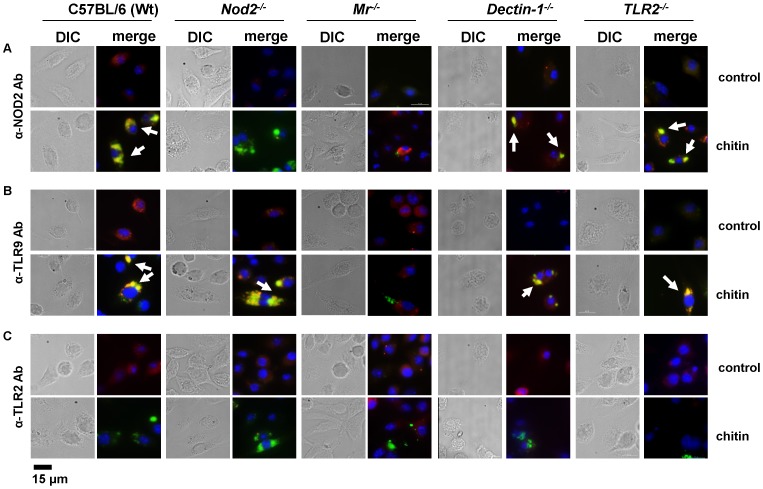
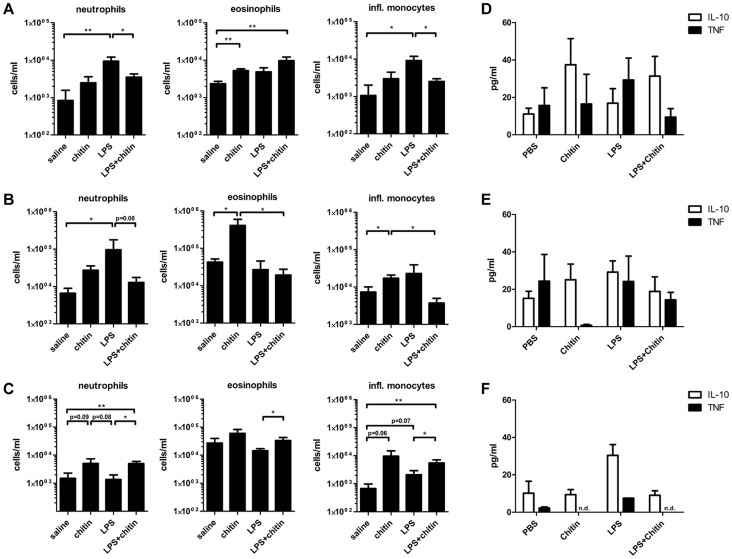
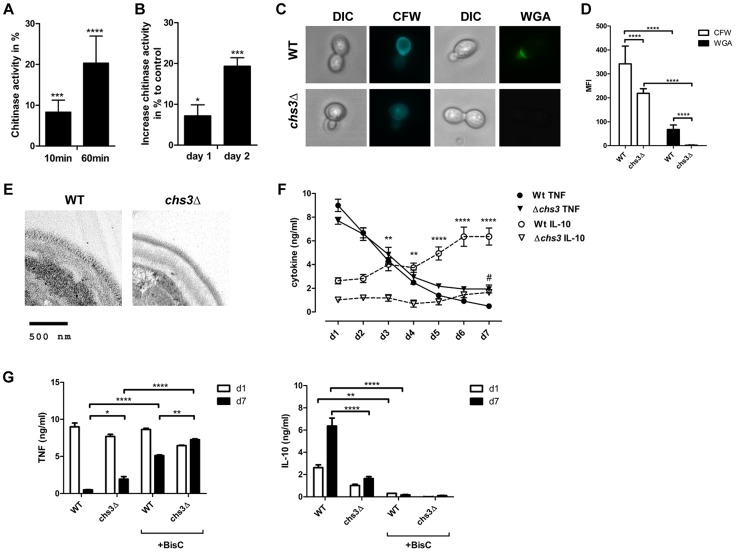
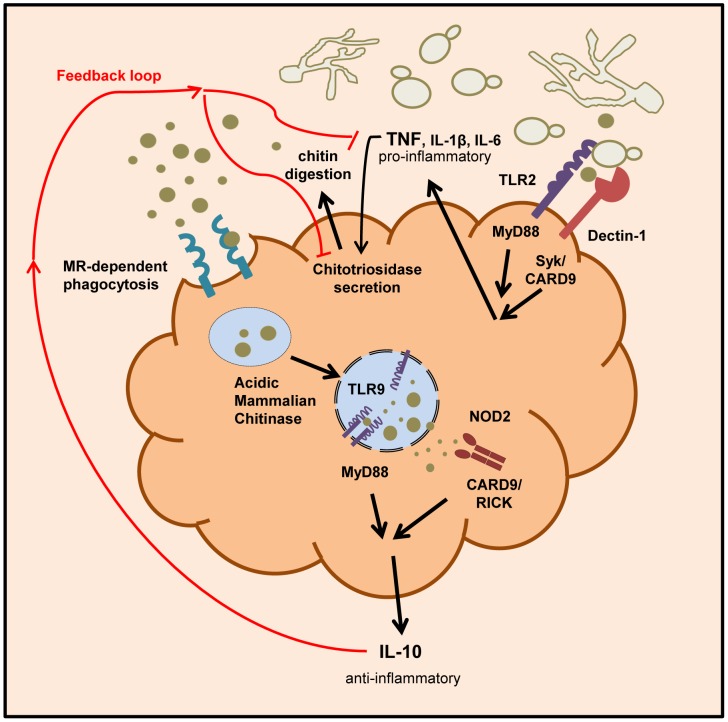
Chitin is an essential structural polysaccharide of fungal pathogens and parasites, but its role in human immune responses remains largely unknown. It is the second most abundant polysaccharide in nature after cellulose and its derivatives today are widely used for medical and industrial purposes. We analysed the immunological properties of purified chitin particles derived from the opportunistic human fungal pathogen Candida albicans, which led to the selective secretion of the anti-inflammatory cytokine IL-10. We identified NOD2, TLR9 and the mannose receptor as essential fungal chitin-recognition receptors for the induction of this response. Chitin reduced LPS-induced inflammation in vivo and may therefore contribute to the resolution of the immune response once the pathogen has been defeated. Fungal chitin also induced eosinophilia in vivo, underpinning its ability to induce asthma. Polymorphisms in the identified chitin receptors, NOD2 and TLR9, predispose individuals to inflammatory conditions and dysregulated expression of chitinases and chitinase-like binding proteins, whose activity is essential to generate IL-10-inducing fungal chitin particles in vitro, have also been linked to inflammatory conditions and asthma. Chitin recognition is therefore critical for immune homeostasis and is likely to have a significant role in infectious and allergic disease.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Chitins and chitosans as immunoadjuvants and non-allergenic drug carriers.Mar Drugs. 2010 Feb 21;8(2):292-312. doi: 10.3390/md8020292. Mar Drugs. 2010. PMID: 20390107 Free PMC article. Review.
-
Chitin, chitinases and chitinase-like proteins in allergic inflammation and tissue remodeling.Yonsei Med J. 2009 Feb 28;50(1):22-30. doi: 10.3349/ymj.2009.50.1.22. Epub 2009 Feb 24. Yonsei Med J. 2009. PMID: 19259344 Free PMC article. Review.
-
Secretory mediators regulate Nod2-induced tolerance in human macrophages.Gastroenterology. 2011 Jan;140(1):231-41. doi: 10.1053/j.gastro.2010.09.009. Epub 2010 Sep 18. Gastroenterology. 2011. PMID: 20854823 Free PMC article.
-
The TLR2-MyD88-NOD2-RIPK2 signalling axis regulates a balanced pro-inflammatory and IL-10-mediated anti-inflammatory cytokine response to Gram-positive cell walls.Cell Microbiol. 2008 Oct;10(10):2067-77. doi: 10.1111/j.1462-5822.2008.01189.x. Epub 2008 Jun 28. Cell Microbiol. 2008. PMID: 18549453 Free PMC article.
-
The fungal ligand chitin directly binds TLR2 and triggers inflammation dependent on oligomer size.EMBO Rep. 2018 Dec;19(12):e46065. doi: 10.15252/embr.201846065. Epub 2018 Oct 18. EMBO Rep. 2018. PMID: 30337494 Free PMC article.
Cited by
-
Induction of Protective Immunity to Cryptococcal Infection in Mice by a Heat-Killed, Chitosan-Deficient Strain of Cryptococcus neoformans.mBio. 2016 May 10;7(3):e00547-16. doi: 10.1128/mBio.00547-16. mBio. 2016. PMID: 27165801 Free PMC article.
-
NOD2 Influences Trajectories of Intestinal Microbiota Recovery After Antibiotic Perturbation.Cell Mol Gastroenterol Hepatol. 2020;10(2):365-389. doi: 10.1016/j.jcmgh.2020.03.008. Epub 2020 Apr 11. Cell Mol Gastroenterol Hepatol. 2020. PMID: 32289499 Free PMC article.
-
Genetic deficiency of NOD2 confers resistance to invasive aspergillosis.Nat Commun. 2018 Jul 6;9(1):2636. doi: 10.1038/s41467-018-04912-3. Nat Commun. 2018. PMID: 29980664 Free PMC article.
-
Pattern recognition receptors in fungal immunity.Semin Cell Dev Biol. 2019 May;89:24-33. doi: 10.1016/j.semcdb.2018.03.003. Epub 2018 Mar 9. Semin Cell Dev Biol. 2019. PMID: 29522806 Free PMC article. Review.
-
Adaptation of Candida albicans to environmental pH induces cell wall remodelling and enhances innate immune recognition.PLoS Pathog. 2017 May 22;13(5):e1006403. doi: 10.1371/journal.ppat.1006403. eCollection 2017 May. PLoS Pathog. 2017. PMID: 28542528 Free PMC article.
References
-
- Araujo AC, Souto-Padron T, de Souza W (1993) Cytochemical localization of carbohydrate residues in microfilariae of Wuchereria bancrofti and Brugia malayi . J Histochem Cytochem 41: 571–578. - PubMed
-
- Fuhrman JA, Piessens WF (1985) Chitin synthesis and sheath morphogenesis in Brugia malayi microfilariae. Mol Biochem Parasitol 17: 93–104. - PubMed
-
- Neville AC, Parry DA, Woodhead-Galloway J (1976) The chitin crystallite in arthropod cuticle. J Cell Sci 21: 73–82. - PubMed
-
- Shahabuddin M, Kaslow DC (1994) Plasmodium: parasite chitinase and its role in malaria transmission. Exp Parasitol 79: 85–88. - PubMed
-
- Brown GD, Denning DW, Gow NA, Levitz SM, Netea MG, et al. (2012) Hidden killers: human fungal infections. Sci Transl Med 4: 165rv113. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 102705/Wellcome Trust/United Kingdom
- 097377/WT_/Wellcome Trust/United Kingdom
- R01 DE017514/DE/NIDCR NIH HHS/United States
- R01 AI101935/AI/NIAID NIH HHS/United States
- 086558/WT_/Wellcome Trust/United Kingdom
- 086827/WT_/Wellcome Trust/United Kingdom
- AR056296/AR/NIAMS NIH HHS/United States
- BB/F00513X/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- Wellcome Trust/United Kingdom
- 101873/Wellcome Trust/United Kingdom
- R01 DE017514-01/DE/NIDCR NIH HHS/United States
- 075470/WT_/Wellcome Trust/United Kingdom
- MR/J008230/1/MRC_/Medical Research Council/United Kingdom
- R37 AI101935/AI/NIAID NIH HHS/United States
- AI101935/AI/NIAID NIH HHS/United States
- 102705/Z/13/Z/Wellcome Trust/United Kingdom
- R01 AR056296/AR/NIAMS NIH HHS/United States
- 080088/WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
