

Review
doi: 10.1083/jcb.201311051.
Review: biogenesis of the multifunctional lipid droplet: lipids, proteins, and sites
Affiliations
- PMID: 24590170
- PMCID: PMC3941045
- DOI: 10.1083/jcb.201311051
Item in Clipboard
Review
Review: biogenesis of the multifunctional lipid droplet: lipids, proteins, and sites
J Cell Biol.
.
Abstract
Lipid droplets (LDs) are ubiquitous dynamic organelles that store and supply lipids in all eukaryotic and some prokaryotic cells for energy metabolism, membrane synthesis, and production of essential lipid-derived molecules. Interest in the organelle's cell biology has exponentially increased over the last decade due to the link between LDs and prevalent human diseases and the discovery of new and unexpected functions of LDs. As a result, there has been significant recent progress toward understanding where and how LDs are formed, and the specific lipid pathways that coordinate LD biogenesis.
Figures
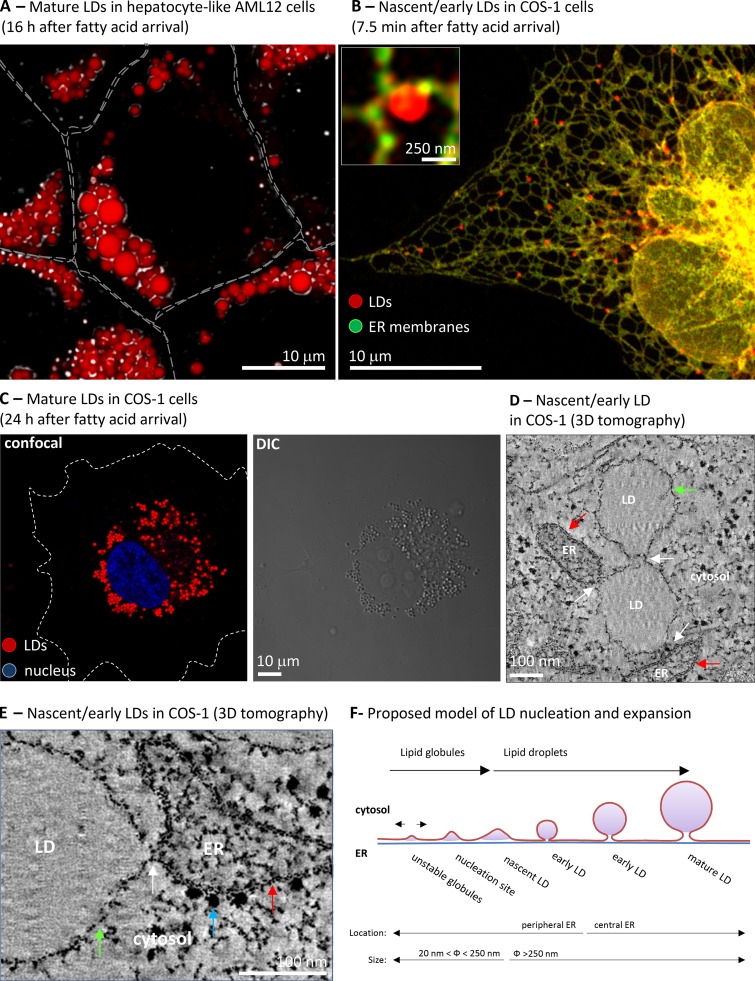
Morphology of LDs. (A) AML-12 cells were treated for 16 h with BSA-bound oleic acid. Cells were fixed and neutral lipids stained with Nile red. The image combines images of confocal microscopy (red) and differential interference contrast images (DIC, black and white). Gray lines indicate the contour of the cells. Image courtesy of Albert Herms. (B) COS-1 cells were cotransfected with OFP-HPos (an LD marker, red) and GFP-HNeu (ER marker, green) and cells were lipid starved during 24 h to reduce the presence of LDs. Then cells were loaded with BSA-bound oleic acid for 7.5 min and the segregation of the markers analyzed by confocal microscopy. The red globular structures correspond to the fatty acid–induced LDs and the inset shows a high magnification image of one of these LDs. The images are from Kassan et al. (2013). (C) Cells treated as in B, but for 24 h, were fixed and neutral lipids stained with Nile red and the nuclei with DAPI (blue). Lipid droplets are visible both by confocal microscopy (left) and DIC (right). White dotted lines indicate the contour of the cell. Image courtesy of Albert Herms. (D and E) LDs induced to form in COS-1 cells were analyzed by electron tomography. The images shows selected low magnification (D) and high magnification (E) sections in which the LD monolayer (green arrow) contacts (white arrow) with the other LDs or the ER bilayer (red arrow), which is recognized by the presence of electron-dense ribosomes (blue arrow). These LDs have an average diameter of 250 nm and thus are nascent or very early LDs. Image from Kassan et al. (2013). (F) Schematic representation of a model of LD biogenesis. Lipid globules produced during triacylglycerol synthesis and deposited between the two ER leaflets of the bilayer that laterally move unless they are nucleated. This nucleation marks the onset of LD biogenesis. From here, LD expansion (sizes are indicated) can be accomplished by recruitment of additional proteins, accumulation of new lipids locally and distally produced, and the synthesis of new phospholipids and lysophospholipids. Finally, the concave nascent LD will be converted into the globular early LD, move into central regions of the cell, and form a mature LD that shows a relatively constant size (Kassan et al., 2013).
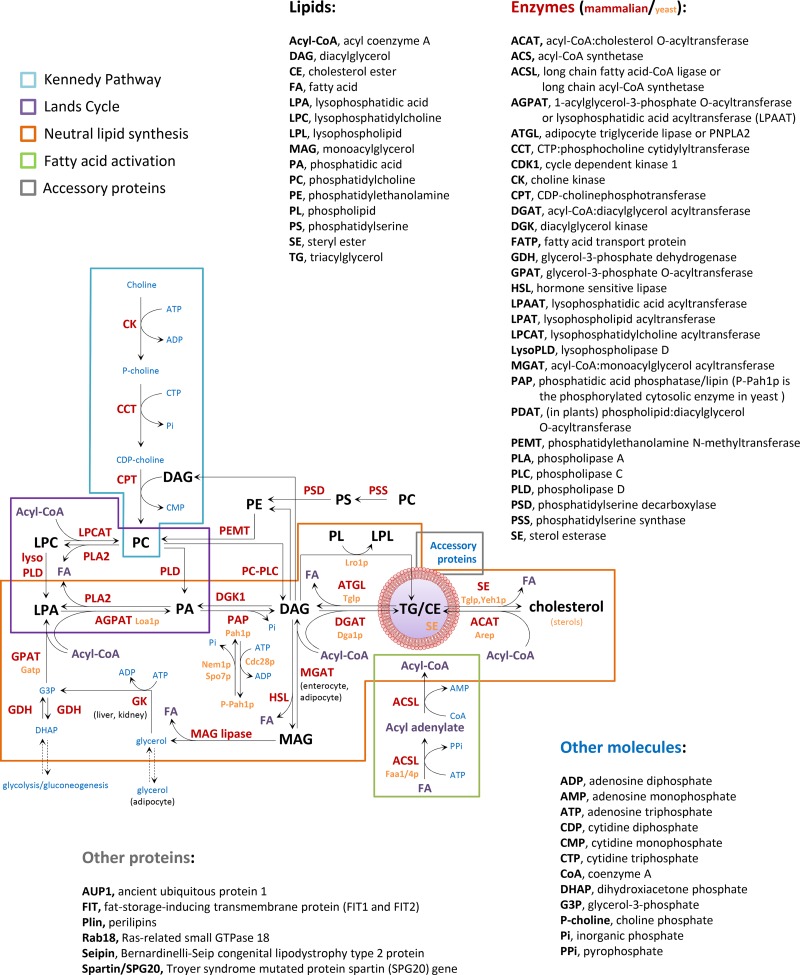
Lipid metabolism and LD formation. LD biogenesis requires coordination of fatty acid activation by acyl-CoA synthetases (green box), de novo synthesis of neutral lipids (orange box), the Lands cycle of phospholipid remodeling (purple box), the Kennedy pathway of phospholipid synthesis (blue box), and the function of accessory proteins (gray box). The enzymes involved in these pathways are indicated with red (mammal) and yellow letters (yeast), the acyl-CoA with purple letters, the lipid intermediates with black letters, and other required molecules with blue letters. See details of lipolysis mechanisms in Zechner et al. (2012).
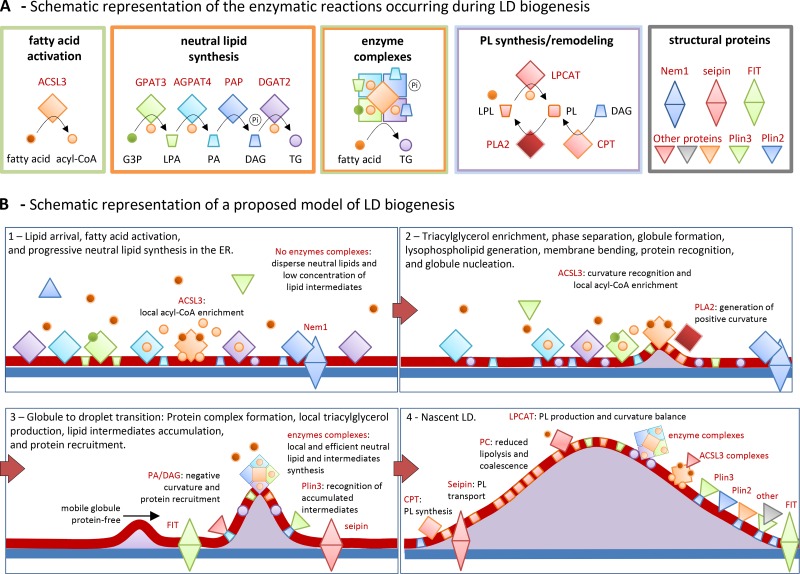
Proposed model of LD biogenesis. (A) Summary of the metabolic pathways detailed in Fig. 2 and occurring on LDs. Small circles and cones indicate lipids, squares enzymes, and triangles accessory protein. The color of each group follows the legend of Fig. 2. The green box illustrates the ACSL3-mediated conversion of the fatty acid into an acyl-CoA. The orange box includes the LD enzymes involved in the local synthesis of triacylglycerol. G3-P, glycerol 3-phosphate; LPA, lysophosphatidic acid; PA, phosphatidic acid; DAG, diacylglycerol; TG, triacylglycerol; Pi, inorganic phosphate. The green/orange box illustrates the protein complexes of neutral lipid synthesis enzymes (orange box) and synthetase activities (green box) proposed to be a cellular mechanism to increase the efficiency of the process by rapidly transferring substrates from one enzyme to the next. The blue and purple box encloses reactions of phospholipid synthesis by the Kennedy pathway (using diacylglycerol as substrate, blue circle) and the Lands cycle of phospholipid remodeling by deacylation and reacylation of phospholipids (PL) and lysophospholipids (LPL). The gray box includes the accessory proteins (single triangles for cytosolic proteins and double triangles for ER-associated proteins). Additional proteins have been added to illustrate in B different mechanisms of protein association with LDs. (B) Proposed model of the stepwise formation of LDs. Shapes and colors follow the legend detailed in A. (B1) Before fatty acid arrival—likely bound to fatty acid transport proteins (not depicted)—neutral lipid synthesis enzymes and ACSL3 are homogenously distributed in the ER tubules. After lipid arrival PAP is recruited to the ER by interaction with Nem1p, perhaps directly to the assembly sites. Lipid intermediates and triacylglycerol are generated along the tubules. However, the enzymes in the proximity of ACSL3 will have availability of at least one required substrate. Finally, a triacylglycerol globule is generated and progressively expands. (B2) At sufficient concentration, the globule of triacylglycerol is not accommodated in the ER membrane, induces phase separation, and it is deposited between the two leaflets of the ER bilayer. The resulting membrane curvature—additionally produced/stabilized by local lysophospholipids or lipid demixing—is recognized by ER proteins with amphipathic helices, such as ACSL3, which stabilize the globule and mark the onset of LD biogenesis. (B3) Transition from a globule into a droplet rapidly occurs by accumulation of ACSL3 that locally increases the acyl-CoA concentration and thus efficiently provides to the neighboring enzymes with a substrate required for many reactions. Neutral synthesis enzymes form complexes at the sites of assembly, illustrated in A. Now triacylglycerol is rapidly produced at the sites of biogenesis, but in addition this pathway locally generates bioactive lipid intermediates. These lipids also determine membrane remodeling and protein recruitment. For example, Plin3 recognizes the accumulation of diacylglycerol and binds to the LD. (B4) The nascent LD rapidly expands into the cytosol by packing mobile globules of triacylglycerol arriving from the rest of the ER (B3, black arrow) and globules locally produced by the enzyme complexes. New phospholipids, to support LD expansion, are generated in neighboring tubules via CPT, and proteins such as seipin and FIT cooperate during the exchange. Conversion of conical lysophospholipids into cylindrical phospholipids, via LPCAT, progressively adjusts the growing structure to the new required curvature and, in addition, the cylindrical phospholipids protect the nascent globule from lipolytic activities and coalescence. Other proteins are anchored to LDs by interaction with lipid species such as phosphatidic acid and diacylglycerol, or by interaction with other LD proteins such as ACSL3. Finally, phosphatidic acid at the base of the structure recruits structural proteins and shapes the concave nascent LD into the globular early LD that will progressively expand and move to central regions of the cell to become a mature LD (Fig. 1 F).
Similar articles
-
Spatial compartmentalization of lipid droplet biogenesis.Biochim Biophys Acta Mol Cell Biol Lipids. 2020 Jan;1865(1):158499. doi: 10.1016/j.bbalip.2019.07.008. Epub 2019 Jul 25. Biochim Biophys Acta Mol Cell Biol Lipids. 2020. PMID: 31352131 Free PMC article. Review.
-
Lipid droplet quantification based on iterative image processing.J Lipid Res. 2019 Jul;60(7):1333-1344. doi: 10.1194/jlr.D092841. Epub 2019 Mar 29. J Lipid Res. 2019. PMID: 30926625 Free PMC article.
-
Acyl-CoA synthetase 3 promotes lipid droplet biogenesis in ER microdomains.J Cell Biol. 2013 Dec 23;203(6):985-1001. doi: 10.1083/jcb.201305142. J Cell Biol. 2013. PMID: 24368806 Free PMC article.
-
Lipid droplets and peroxisomes: key players in cellular lipid homeostasis or a matter of fat--store 'em up or burn 'em down.Genetics. 2013 Jan;193(1):1-50. doi: 10.1534/genetics.112.143362. Genetics. 2013. PMID: 23275493 Free PMC article. Review.
-
Lipid droplet growth: regulation of a dynamic organelle.Curr Opin Cell Biol. 2017 Aug;47:9-15. doi: 10.1016/j.ceb.2017.02.002. Epub 2017 Feb 21. Curr Opin Cell Biol. 2017. PMID: 28231490 Review.
Cited by
-
Expanding roles for lipid droplets.Curr Biol. 2015 Jun 1;25(11):R470-81. doi: 10.1016/j.cub.2015.04.004. Curr Biol. 2015. PMID: 26035793 Free PMC article. Review.
-
Spatial control of lipid droplet proteins by the ERAD ubiquitin ligase Doa10.EMBO J. 2016 Aug 1;35(15):1644-55. doi: 10.15252/embj.201593106. Epub 2016 Jun 29. EMBO J. 2016. PMID: 27357570 Free PMC article.
-
A transcriptomic examination of encased rotifer embryos reveals the developmental trajectory leading to long-term dormancy; are they "animal seeds"?BMC Genomics. 2024 Jan 27;25(1):119. doi: 10.1186/s12864-024-09961-1. BMC Genomics. 2024. PMID: 38281016 Free PMC article.
-
As the fat flies: The dynamic lipid droplets of Drosophila embryos.Biochim Biophys Acta. 2015 Sep;1851(9):1156-85. doi: 10.1016/j.bbalip.2015.04.002. Epub 2015 Apr 13. Biochim Biophys Acta. 2015. PMID: 25882628 Free PMC article. Review.
-
Septin 9 induces lipid droplets growth by a phosphatidylinositol-5-phosphate and microtubule-dependent mechanism hijacked by HCV.Nat Commun. 2016 Jul 15;7:12203. doi: 10.1038/ncomms12203. Nat Commun. 2016. PMID: 27417143 Free PMC article.
References
-
- Ayciriex S., Le Guédard M., Camougrand N., Velours G., Schoene M., Leone S., Wattelet-Boyer V., Dupuy J.W., Shevchenko A., Schmitter J.M., et al. 2012. YPR139c/LOA1 encodes a novel lysophosphatidic acid acyltransferase associated with lipid droplets and involved in TAG homeostasis. Mol. Biol. Cell. 23:233–246 10.1091/mbc.E11-07-0650 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
