

Structural basis for Klf4 recognition of methylated DNA
- PMID: 24520114
- PMCID: PMC4005678
- DOI: 10.1093/nar/gku134
Structural basis for Klf4 recognition of methylated DNA
Abstract
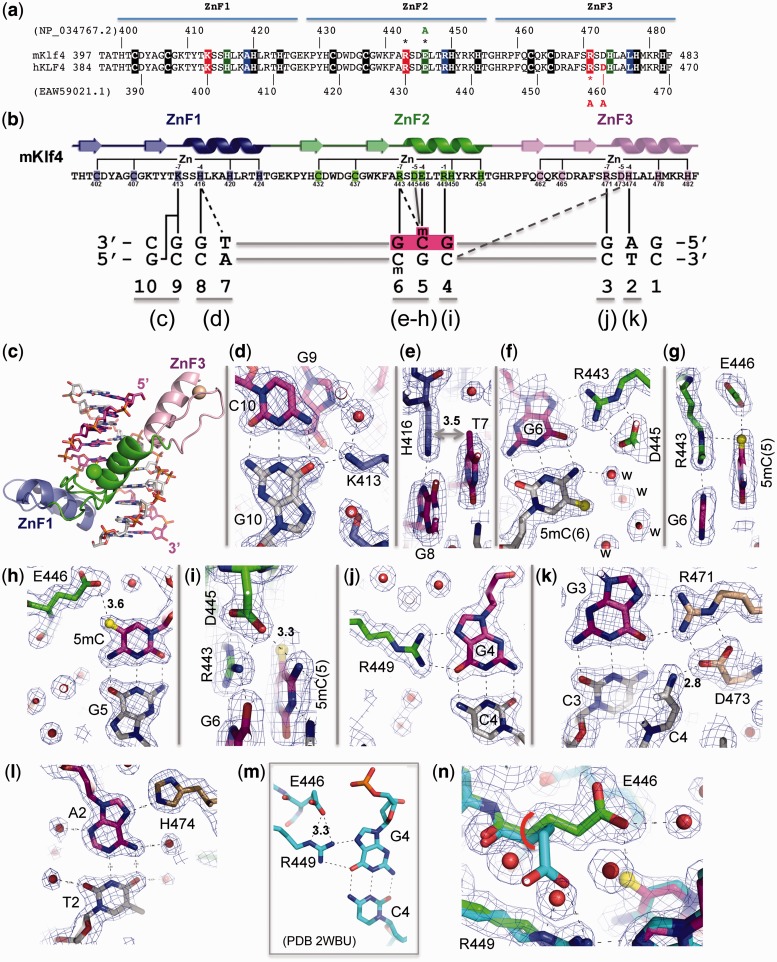
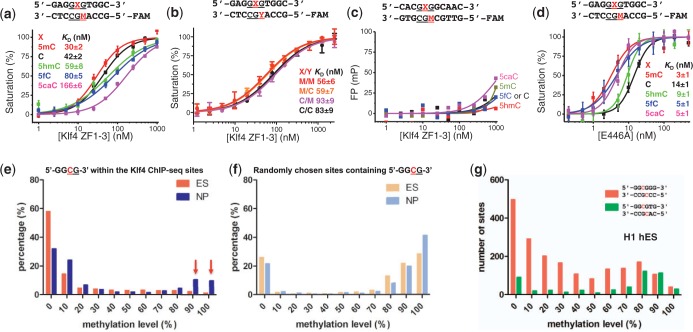
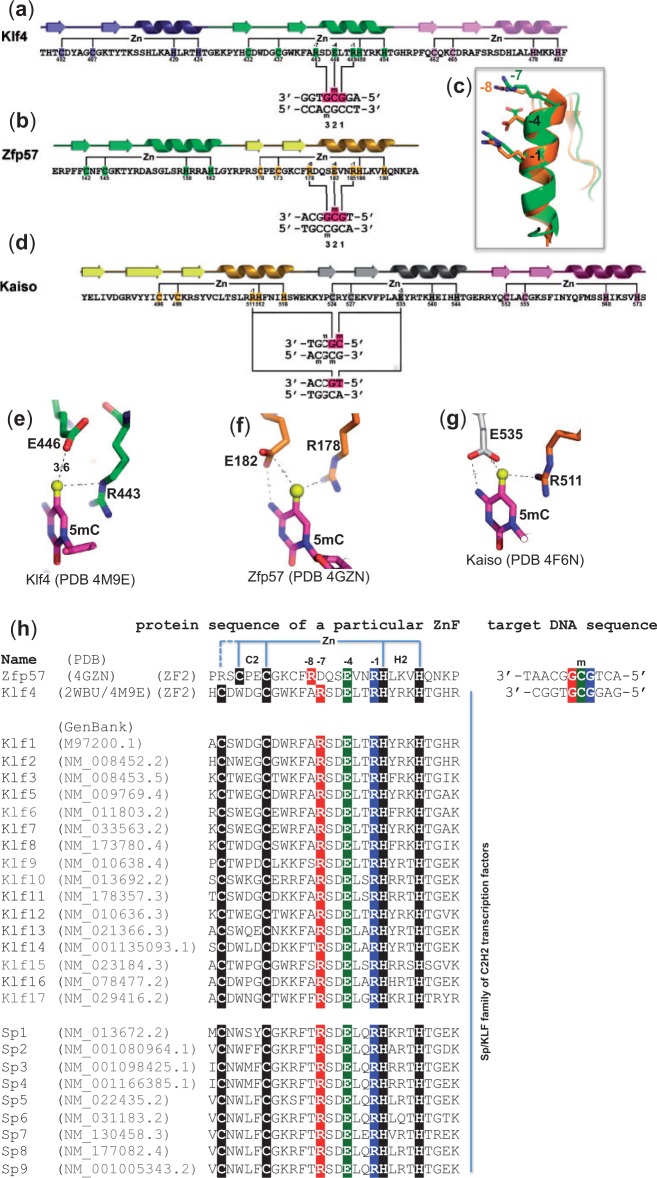
Transcription factor Krüppel-like factor 4 (Klf4), one of the factors directing cellular reprogramming, recognizes the CpG dinucleotide (whether methylated or unmodified) within a specific G/C-rich sequence. The binding affinity of the mouse Klf4 DNA-binding domain for methylated DNA is only slightly stronger than that for an unmodified oligonucleotide. The structure of the C-terminal three Krüppel-like zinc fingers (ZnFs) of mouse Klf4, in complex with fully methylated DNA, was determined at 1.85 Å resolution. An arginine and a glutamate interact with the methyl group. By comparison with two other recently characterized structures of ZnF protein complexes with methylated DNA, we propose a common principle of recognition of methylated CpG by C2H2 ZnF proteins, which involves a spatially conserved Arg-Glu pair.
Figures
Similar articles
-
Analysis of KLF4 regulated genes in cancer cells reveals a role of DNA methylation in promoter- enhancer interactions.Epigenetics. 2018;13(7):751-768. doi: 10.1080/15592294.2018.1504592. Epub 2018 Aug 25. Epigenetics. 2018. PMID: 30058478 Free PMC article.
-
Distinctive Klf4 mutants determine preference for DNA methylation status.Nucleic Acids Res. 2016 Dec 1;44(21):10177-10185. doi: 10.1093/nar/gkw774. Epub 2016 Sep 4. Nucleic Acids Res. 2016. PMID: 27596594 Free PMC article.
-
A common mode of recognition for methylated CpG.Trends Biochem Sci. 2013 Apr;38(4):177-83. doi: 10.1016/j.tibs.2012.12.005. Epub 2013 Jan 23. Trends Biochem Sci. 2013. PMID: 23352388 Free PMC article. Review.
-
An atomic model of Zfp57 recognition of CpG methylation within a specific DNA sequence.Genes Dev. 2012 Nov 1;26(21):2374-9. doi: 10.1101/gad.202200.112. Epub 2012 Oct 11. Genes Dev. 2012. PMID: 23059534 Free PMC article.
-
On how mammalian transcription factors recognize methylated DNA.Epigenetics. 2013 Feb;8(2):131-7. doi: 10.4161/epi.23632. Epub 2013 Jan 16. Epigenetics. 2013. PMID: 23324617 Free PMC article. Review.
Cited by
-
Analysis of KLF4 regulated genes in cancer cells reveals a role of DNA methylation in promoter- enhancer interactions.Epigenetics. 2018;13(7):751-768. doi: 10.1080/15592294.2018.1504592. Epub 2018 Aug 25. Epigenetics. 2018. PMID: 30058478 Free PMC article.
-
DNA methylation: old dog, new tricks?Nat Struct Mol Biol. 2014 Nov;21(11):949-54. doi: 10.1038/nsmb.2910. Nat Struct Mol Biol. 2014. PMID: 25372310
-
Liquid condensation of reprogramming factor KLF4 with DNA provides a mechanism for chromatin organization.Nat Commun. 2021 Sep 22;12(1):5579. doi: 10.1038/s41467-021-25761-7. Nat Commun. 2021. PMID: 34552088 Free PMC article.
-
Detecting and interpreting DNA methylation marks.Curr Opin Struct Biol. 2018 Dec;53:88-99. doi: 10.1016/j.sbi.2018.06.004. Epub 2018 Jul 19. Curr Opin Struct Biol. 2018. PMID: 30031306 Free PMC article. Review.
-
Long noncoding RNA TUG1 regulates smooth muscle cell differentiation via KLF4-myocardin axis.Am J Physiol Cell Physiol. 2023 Oct 1;325(4):C940-C950. doi: 10.1152/ajpcell.00275.2023. Epub 2023 Aug 29. Am J Physiol Cell Physiol. 2023. PMID: 37642238 Free PMC article.
References
-
- Bestor T, Laudano A, Mattaliano R, Ingram V. Cloning and sequencing of a cDNA encoding DNA methyltransferase of mouse cells. The carboxyl-terminal domain of the mammalian enzymes is related to bacterial restriction methyltransferases. J. Mol. Biol. 1988;203:971–983. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
