

Selective mRNA sequestration by OLIGOURIDYLATE-BINDING PROTEIN 1 contributes to translational control during hypoxia in Arabidopsis
- PMID: 24469793
- PMCID: PMC3926019
- DOI: 10.1073/pnas.1314851111
Selective mRNA sequestration by OLIGOURIDYLATE-BINDING PROTEIN 1 contributes to translational control during hypoxia in Arabidopsis
Abstract
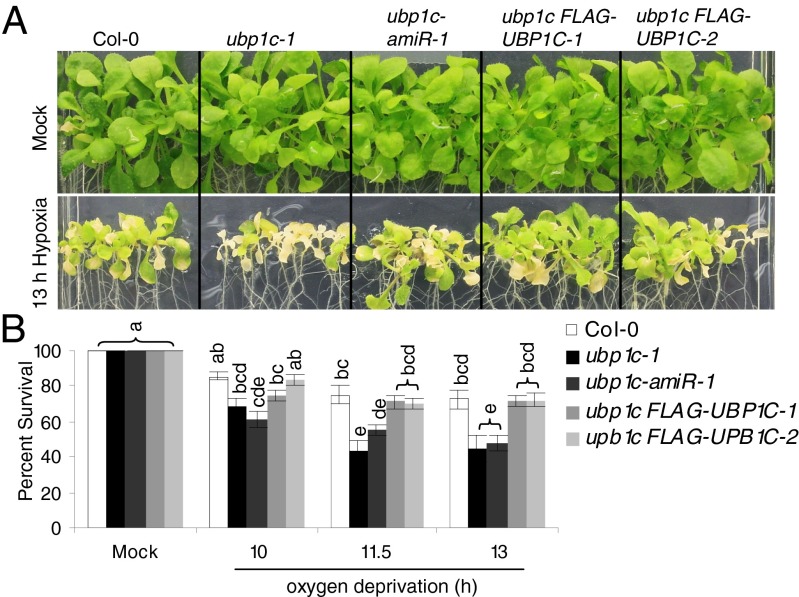
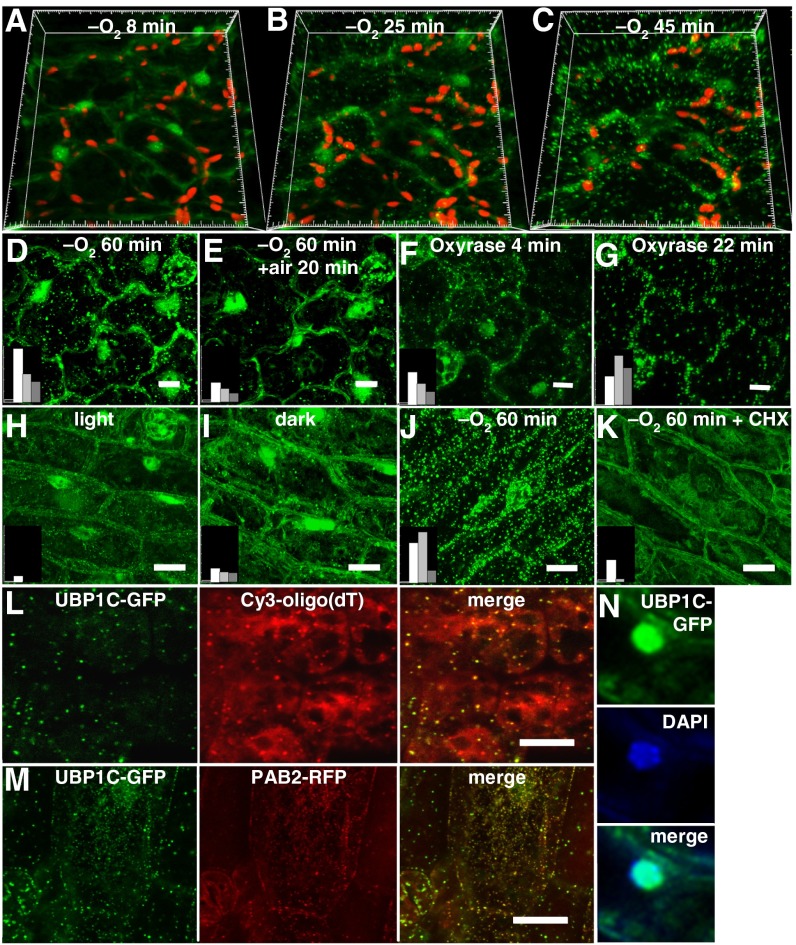
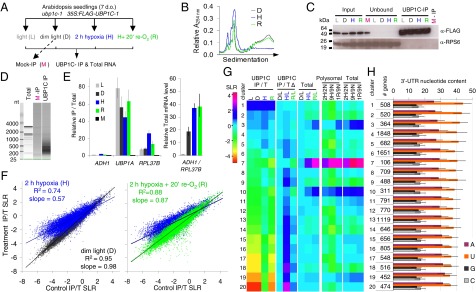
Low oxygen stress dynamically regulates the translation of cellular mRNAs as a means of energy conservation in seedlings of Arabidopsis thaliana. Most of the highly hypoxia-induced mRNAs are recruited to polysomes and actively translated, whereas other cellular mRNAs become translationally inactive and are either targeted for stabilization or degradation. Here we identify the involvement of OLIGOURIDYLATE BINDING PROTEIN 1 (UBP1), a triple RNA Recognition Motif protein, in dynamic and reversible aggregation of translationally repressed mRNAs during hypoxia. Mutation or down-regulation of UBP1C interferes with seedling establishment and reduces survival of low oxygen stress. By use of messenger ribonucleoprotein (mRNP) immunopurification, we show that UBP1C constitutively binds a subpopulation of mRNAs characterized by uracil-rich 3'-untranslated regions under normoxic conditions. During hypoxia, UBP1C association with non-uracil-rich mRNAs is enhanced concomitant with its aggregation into microscopically visible cytoplasmic foci, referred to as UBP1 stress granules (SGs). This UBP1C-mRNA association occurs as global levels of protein synthesis decline. Upon reoxygenation, rapid UBP1 SG disaggregation coincides with the return of the stabilized mRNAs to polysomes. The mRNAs that are highly induced and translated during hypoxia largely circumvent UBP1C sequestration. Thus, UBP1 is established as a component of dynamically assembled cytoplasmic mRNPs that sequester mRNAs that are poorly translated during a transient low energy stress.
Keywords: RNA-binding; TIA-1; posttranscriptional; ribosome.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Genome-wide analysis of transcript abundance and translation in Arabidopsis seedlings subjected to oxygen deprivation.Ann Bot. 2005 Sep;96(4):647-60. doi: 10.1093/aob/mci217. Epub 2005 Aug 4. Ann Bot. 2005. PMID: 16081496 Free PMC article.
-
Plant stress granules and mRNA processing bodies are distinct from heat stress granules.Plant J. 2008 Nov;56(4):517-30. doi: 10.1111/j.1365-313X.2008.03623.x. Epub 2008 Aug 6. Plant J. 2008. PMID: 18643965
-
Translational dynamics revealed by genome-wide profiling of ribosome footprints in Arabidopsis.Proc Natl Acad Sci U S A. 2014 Jan 7;111(1):E203-12. doi: 10.1073/pnas.1317811111. Epub 2013 Dec 23. Proc Natl Acad Sci U S A. 2014. PMID: 24367078 Free PMC article.
-
Stress Granules and Processing Bodies in Translational Control.Cold Spring Harb Perspect Biol. 2019 May 1;11(5):a032813. doi: 10.1101/cshperspect.a032813. Cold Spring Harb Perspect Biol. 2019. PMID: 30082464 Free PMC article. Review.
-
P-bodies and stress granules: possible roles in the control of translation and mRNA degradation.Cold Spring Harb Perspect Biol. 2012 Sep 1;4(9):a012286. doi: 10.1101/cshperspect.a012286. Cold Spring Harb Perspect Biol. 2012. PMID: 22763747 Free PMC article. Review.
Cited by
-
Plant Long Noncoding RNAs: New Players in the Field of Post-Transcriptional Regulations.Noncoding RNA. 2021 Feb 17;7(1):12. doi: 10.3390/ncrna7010012. Noncoding RNA. 2021. PMID: 33671131 Free PMC article. Review.
-
Reprogramming of Root Cells during Nitrogen-Fixing Symbiosis Involves Dynamic Polysome Association of Coding and Noncoding RNAs.Plant Cell. 2020 Feb;32(2):352-373. doi: 10.1105/tpc.19.00647. Epub 2019 Nov 20. Plant Cell. 2020. PMID: 31748328 Free PMC article.
-
Ethylene-Regulated Glutamate Dehydrogenase Fine-Tunes Metabolism during Anoxia-Reoxygenation.Plant Physiol. 2016 Nov;172(3):1548-1562. doi: 10.1104/pp.16.00985. Epub 2016 Sep 27. Plant Physiol. 2016. PMID: 27677986 Free PMC article.
-
Dynamics and Functions of Stress Granules and Processing Bodies in Plants.Plants (Basel). 2020 Aug 30;9(9):1122. doi: 10.3390/plants9091122. Plants (Basel). 2020. PMID: 32872650 Free PMC article. Review.
-
Targeted Endoplasmic Reticulum Localization of Storage Protein mRNAs Requires the RNA-Binding Protein RBP-L.Plant Physiol. 2019 Mar;179(3):1111-1131. doi: 10.1104/pp.18.01434. Epub 2019 Jan 18. Plant Physiol. 2019. PMID: 30659066 Free PMC article.
References
-
- Branco-Price C, Kaiser KA, Jang CJH, Larive CK, Bailey-Serres J. Selective mRNA translation coordinates energetic and metabolic adjustments to cellular oxygen deprivation and reoxygenation in Arabidopsis thaliana. Plant J. 2008;56(5):743–755. - PubMed
-
- Ivanov N. Stress granules: RNP-containing cytoplasmic bodies arising in stress: Structure and mechanism of organization. Mol Biol. 2006;40(6):844–850. - PubMed
-
- Anderson P, Kedersha N. RNA granules: Post-transcriptional and epigenetic modulators of gene expression. Nat Rev Mol Cell Biol. 2009;10(6):430–436. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
