

The progression of peroxisomal degradation through autophagy requires peroxisomal division
- PMID: 24451165
- PMCID: PMC4091152
- DOI: 10.4161/auto.27852
The progression of peroxisomal degradation through autophagy requires peroxisomal division
Abstract
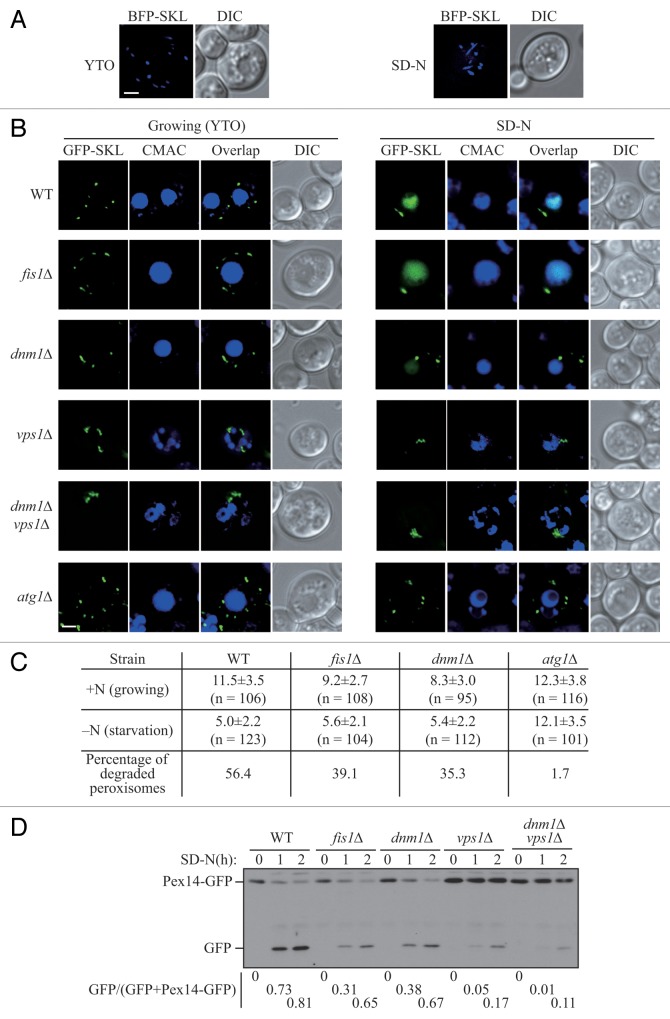
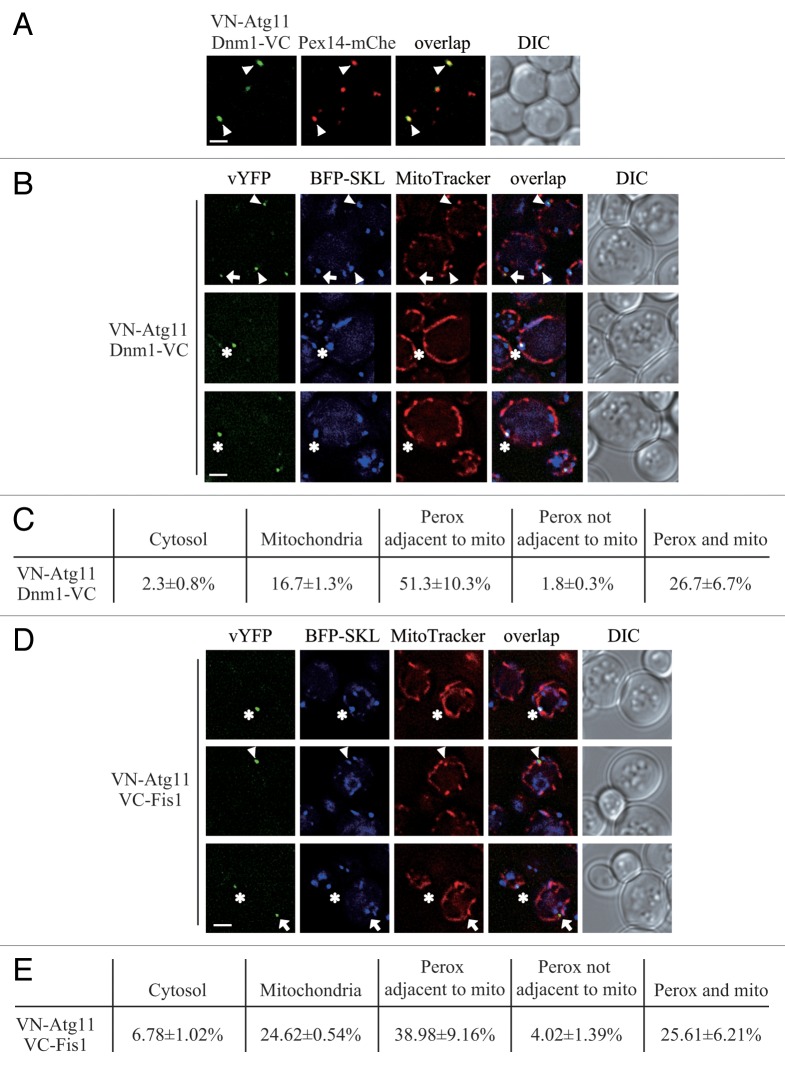
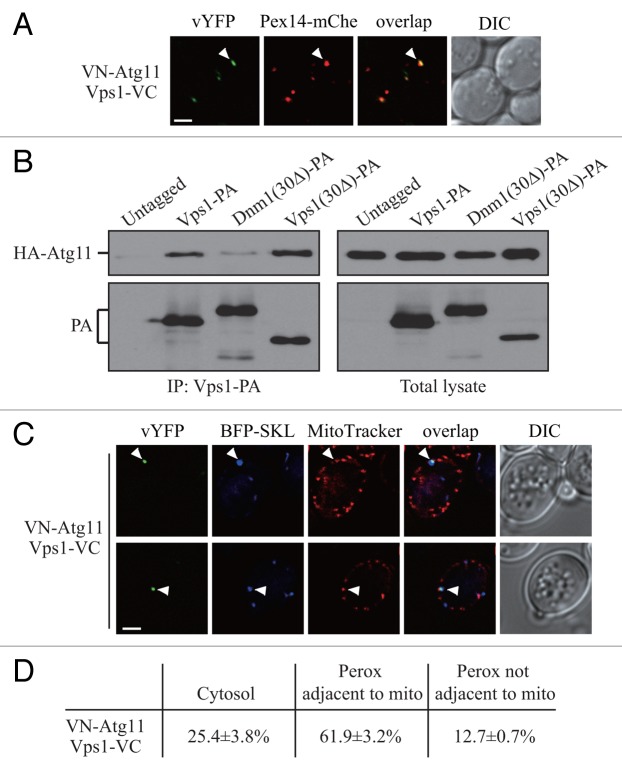
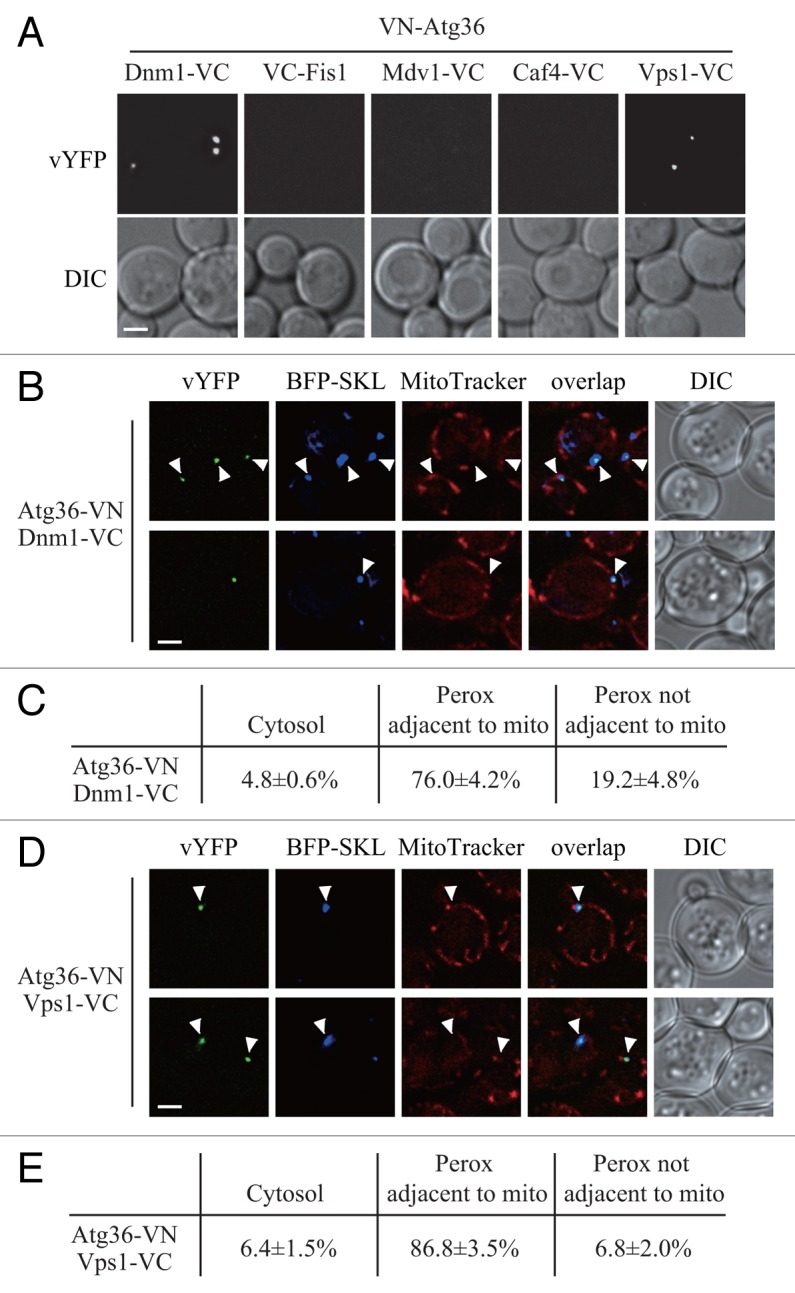
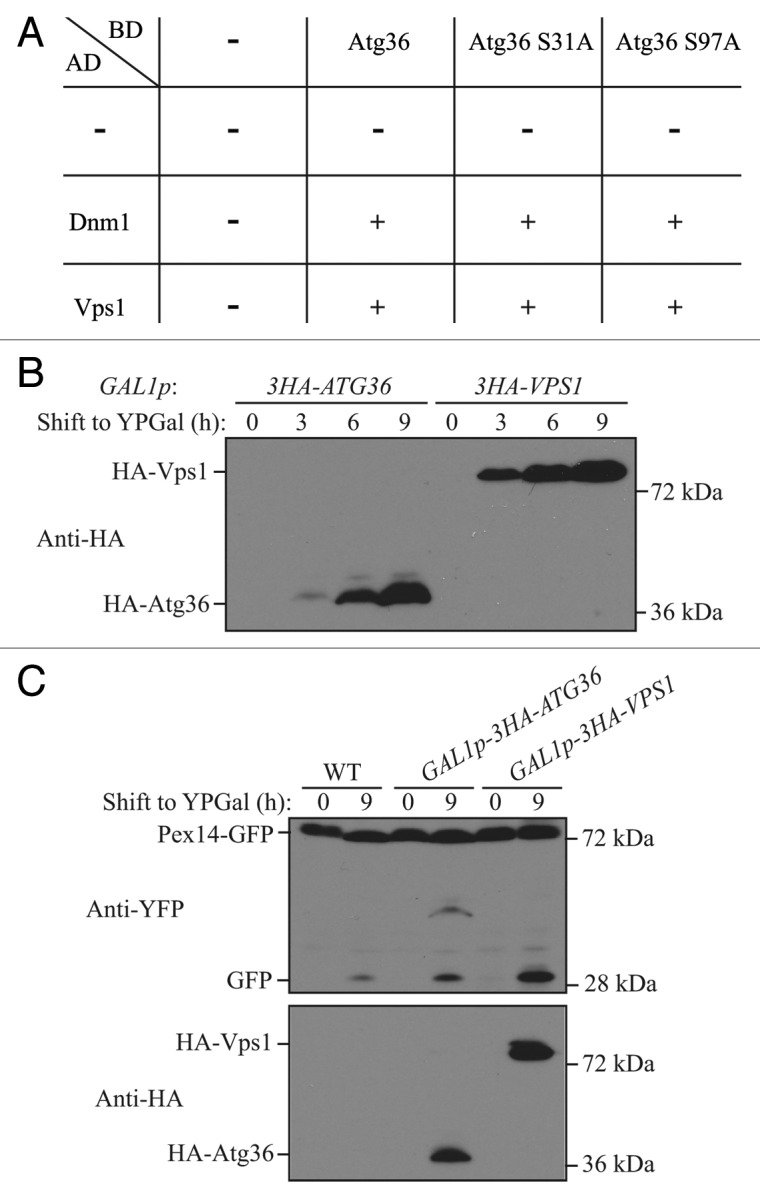
Peroxisomes are highly dynamic organelles that have multiple functions in cellular metabolism. To adapt the intracellular conditions to the changing extracellular environment, peroxisomes undergo constitutive segregation and degradation. The segregation of peroxisomes is mediated by 2 dynamin-related GTPases, Dnm1 and Vps1, whereas, the degradation of peroxisomes is accomplished through pexophagy, a selective type of autophagy. During pexophagy, the size of the organelle is always a challenging factor for the efficiency of engulfment by the sequestering compartment, the phagophore, which implies a potential role for peroxisomal fission in the degradation process, similar to the situation with selective mitochondria degradation. In this study, we report that peroxisomal fission is indeed critical for the efficient elimination of the organelle. When pexophagy is induced, both Dnm1 and Vps1 are recruited to the degrading peroxisomes through interactions with Atg11 and Atg36. In addition, we found that specific peroxisomal fission, which is only needed for pexophagy, occurs at mitochondria-peroxisome contact sites.
Keywords: pexophagy; phagophore; stress; vacuole; yeast.
Figures
Similar articles
-
Pex3-anchored Atg36 tags peroxisomes for degradation in Saccharomyces cerevisiae.EMBO J. 2012 Jun 29;31(13):2852-68. doi: 10.1038/emboj.2012.151. Epub 2012 May 29. EMBO J. 2012. PMID: 22643220 Free PMC article.
-
ER-mitochondria contacts are required for pexophagy in Saccharomyces cerevisiae.Contact (Thousand Oaks). 2018 Jan-Dec;2:10.1177/2515256418821584. doi: 10.1177/2515256418821584. Epub 2019 Jan 8. Contact (Thousand Oaks). 2018. PMID: 30859155 Free PMC article.
-
Peroxisomal Pex3 activates selective autophagy of peroxisomes via interaction with the pexophagy receptor Atg30.J Biol Chem. 2015 Mar 27;290(13):8623-31. doi: 10.1074/jbc.M114.619338. Epub 2015 Feb 18. J Biol Chem. 2015. PMID: 25694426 Free PMC article.
-
Pexophagy: Molecular Mechanisms and Implications for Health and Diseases.Mol Cells. 2018 Jan 31;41(1):55-64. doi: 10.14348/molcells.2018.2245. Epub 2018 Jan 23. Mol Cells. 2018. PMID: 29370694 Free PMC article. Review.
-
Peroxisome homeostasis: Mechanisms of division and selective degradation of peroxisomes in mammals.Biochim Biophys Acta. 2016 May;1863(5):984-91. doi: 10.1016/j.bbamcr.2015.09.032. Epub 2015 Oct 3. Biochim Biophys Acta. 2016. PMID: 26434997 Review.
Cited by
-
Target acquired: Selective autophagy in cardiometabolic disease.Sci Signal. 2017 Feb 28;10(468):eaag2298. doi: 10.1126/scisignal.aag2298. Sci Signal. 2017. PMID: 28246200 Free PMC article. Review.
-
Convergent and divergent mechanisms of peroxisomal and mitochondrial division.J Cell Biol. 2023 Sep 4;222(9):e202304076. doi: 10.1083/jcb.202304076. Epub 2023 Aug 2. J Cell Biol. 2023. PMID: 37530713 Free PMC article.
-
The different facets of organelle interplay-an overview of organelle interactions.Front Cell Dev Biol. 2015 Sep 25;3:56. doi: 10.3389/fcell.2015.00056. eCollection 2015. Front Cell Dev Biol. 2015. PMID: 26442263 Free PMC article. Review.
-
Redox regulated peroxisome homeostasis.Redox Biol. 2015;4:104-8. doi: 10.1016/j.redox.2014.12.006. Epub 2014 Dec 18. Redox Biol. 2015. PMID: 25545794 Free PMC article. Review.
-
Autophagy-related proteins are functionally active in human spermatozoa and may be involved in the regulation of cell survival and motility.Sci Rep. 2016 Sep 16;6:33647. doi: 10.1038/srep33647. Sci Rep. 2016. PMID: 27633131 Free PMC article.
References
-
- Hutchins MU, Veenhuis M, Klionsky DJ. Peroxisome degradation in Saccharomyces cerevisiae is dependent on machinery of macroautophagy and the Cvt pathway. J Cell Sci. 1999;112:4079–87. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases