

The yeast p5 type ATPase, spf1, regulates manganese transport into the endoplasmic reticulum
- PMID: 24392018
- PMCID: PMC3877380
- DOI: 10.1371/journal.pone.0085519
The yeast p5 type ATPase, spf1, regulates manganese transport into the endoplasmic reticulum
Abstract
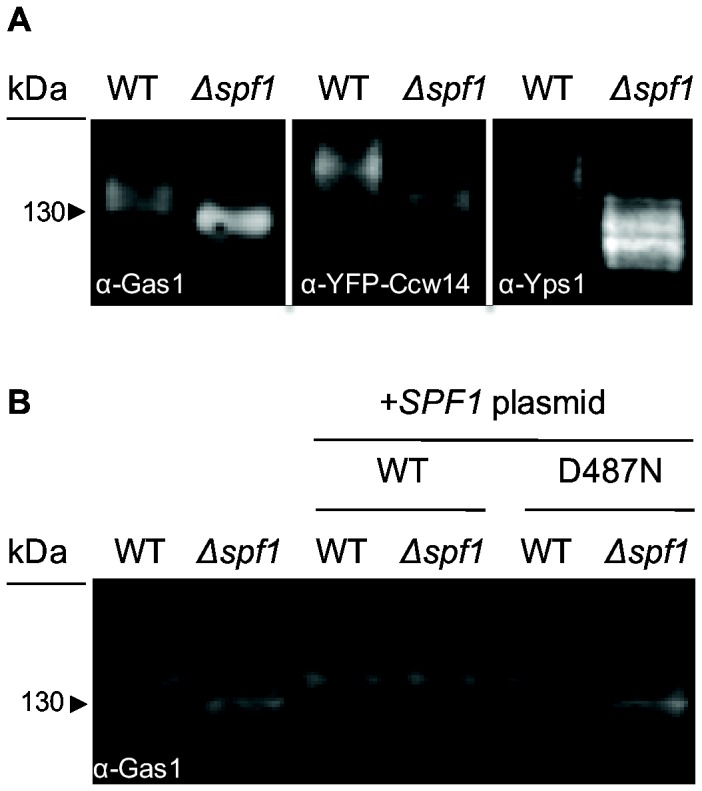
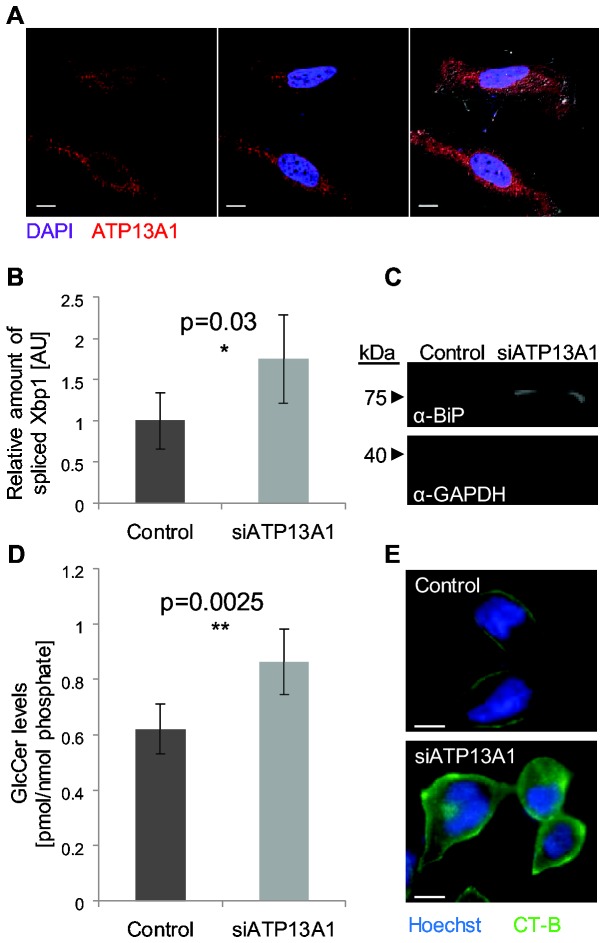
The endoplasmic reticulum (ER) is a large, multifunctional and essential organelle. Despite intense research, the function of more than a third of ER proteins remains unknown even in the well-studied model organism Saccharomyces cerevisiae. One such protein is Spf1, which is a highly conserved, ER localized, putative P-type ATPase. Deletion of SPF1 causes a wide variety of phenotypes including severe ER stress suggesting that this protein is essential for the normal function of the ER. The closest homologue of Spf1 is the vacuolar P-type ATPase Ypk9 that influences Mn(2+) homeostasis. However in vitro reconstitution assays with Spf1 have not yielded insight into its transport specificity. Here we took an in vivo approach to detect the direct and indirect effects of deleting SPF1. We found a specific reduction in the luminal concentration of Mn(2+) in ∆spf1 cells and an increase following it's overexpression. In agreement with the observed loss of luminal Mn(2+) we could observe concurrent reduction in many Mn(2+)-related process in the ER lumen. Conversely, cytosolic Mn(2+)-dependent processes were increased. Together, these data support a role for Spf1p in Mn(2+) transport in the cell. We also demonstrate that the human sequence homologue, ATP13A1, is a functionally conserved orthologue. Since ATP13A1 is highly expressed in developing neuronal tissues and in the brain, this should help in the study of Mn(2+)-dependent neurological disorders.
Conflict of interest statement
Figures
Similar articles
-
The P5A ATPase Spf1p is stimulated by phosphatidylinositol 4-phosphate and influences cellular sterol homeostasis.Mol Biol Cell. 2019 Apr 15;30(9):1069-1084. doi: 10.1091/mbc.E18-06-0365. Epub 2019 Feb 20. Mol Biol Cell. 2019. PMID: 30785834 Free PMC article.
-
Cooperative function of the CHD5-like protein Mdm39p with a P-type ATPase Spf1p in the maintenance of ER homeostasis in Saccharomyces cerevisiae.Mol Genet Genomics. 2005 Jul;273(6):497-506. doi: 10.1007/s00438-005-1153-6. Epub 2005 May 21. Mol Genet Genomics. 2005. PMID: 15909163
-
Manganese redistribution by calcium-stimulated vesicle trafficking bypasses the need for P-type ATPase function.J Biol Chem. 2015 Apr 10;290(15):9335-47. doi: 10.1074/jbc.M114.616334. Epub 2015 Feb 20. J Biol Chem. 2015. PMID: 25713143 Free PMC article.
-
Towards defining the substrate of orphan P5A-ATPases.Biochim Biophys Acta. 2015 Mar;1850(3):524-35. doi: 10.1016/j.bbagen.2014.05.008. Epub 2014 May 14. Biochim Biophys Acta. 2015. PMID: 24836520 Review.
-
Cryo-EM insights into tail-anchored membrane protein biogenesis in eukaryotes.Curr Opin Struct Biol. 2022 Aug;75:102428. doi: 10.1016/j.sbi.2022.102428. Epub 2022 Jul 16. Curr Opin Struct Biol. 2022. PMID: 35850079 Review.
Cited by
-
Effect of cadmium on essential metals and their impact on lipid metabolism in Saccharomyces cerevisiae.Cell Stress Chaperones. 2020 Jan;25(1):19-33. doi: 10.1007/s12192-019-01058-z. Epub 2019 Dec 10. Cell Stress Chaperones. 2020. PMID: 31823289 Free PMC article. Review.
-
The P5-type ATPase ATP13A1 modulates major histocompatibility complex I-related protein 1 (MR1)-mediated antigen presentation.J Biol Chem. 2022 Feb;298(2):101542. doi: 10.1016/j.jbc.2021.101542. Epub 2021 Dec 27. J Biol Chem. 2022. PMID: 34968463 Free PMC article.
-
Reduction of the P5A-ATPase Spf1p phosphoenzyme by a Ca2+-dependent phosphatase.PLoS One. 2020 Apr 30;15(4):e0232476. doi: 10.1371/journal.pone.0232476. eCollection 2020. PLoS One. 2020. PMID: 32353073 Free PMC article.
-
Cancer associated mutations in Sec61γ alter the permeability of the ER translocase.PLoS Genet. 2021 Aug 30;17(8):e1009780. doi: 10.1371/journal.pgen.1009780. eCollection 2021 Aug. PLoS Genet. 2021. PMID: 34460824 Free PMC article.
-
Impaired XK recycling for importing manganese underlies striatal vulnerability in Huntington's disease.J Cell Biol. 2022 Oct 3;221(10):e202112073. doi: 10.1083/jcb.202112073. Epub 2022 Sep 13. J Cell Biol. 2022. PMID: 36099524 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
