

Human 4E-T represses translation of bound mRNAs and enhances microRNA-mediated silencing
- PMID: 24335285
- PMCID: PMC3950672
- DOI: 10.1093/nar/gkt1265
Human 4E-T represses translation of bound mRNAs and enhances microRNA-mediated silencing
Abstract
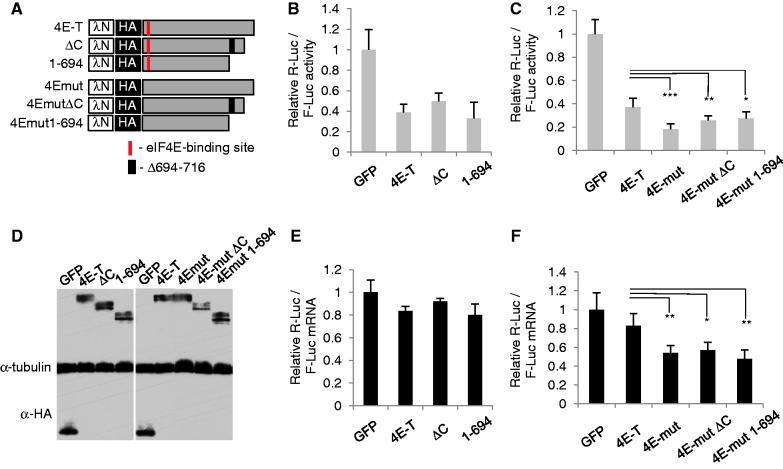
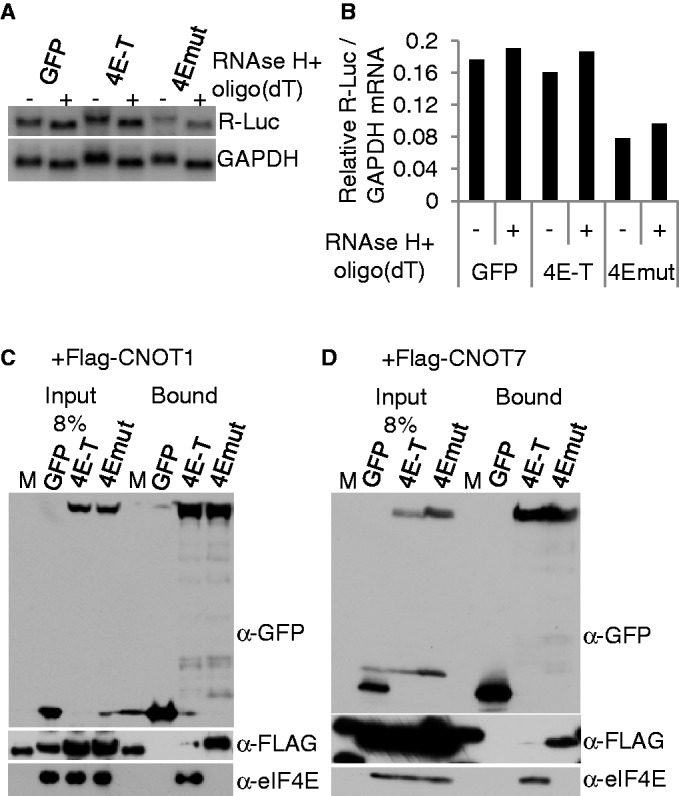
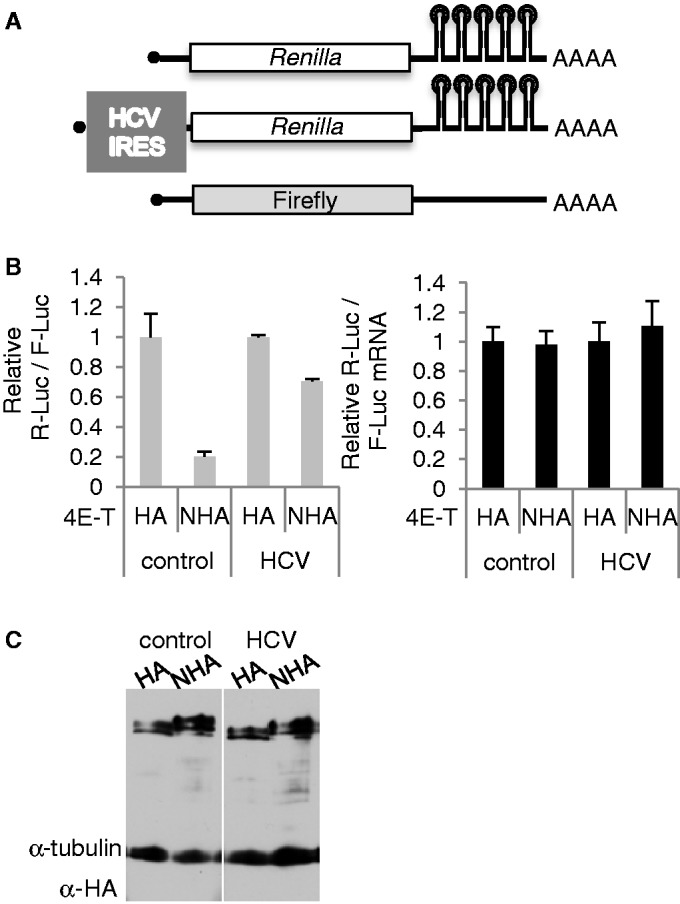
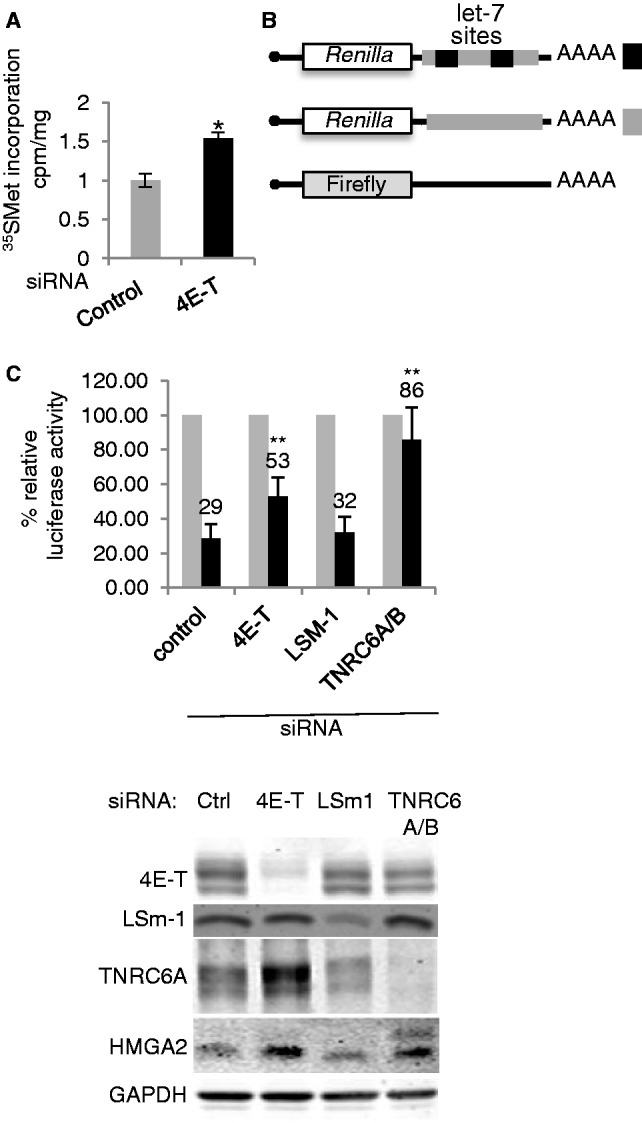
A key player in translation initiation is eIF4E, the mRNA 5' cap-binding protein. 4E-Transporter (4E-T) is a recently characterized eIF4E-binding protein, which regulates specific mRNAs in several developmental model systems. Here, we first investigated the role of its enrichment in P-bodies and eIF4E-binding in translational regulation in mammalian cells. Identification of the conserved C-terminal sequences that target 4E-T to P-bodies was enabled by comparison of vertebrate proteins with homologues in Drosophila (Cup and CG32016) and Caenorhabditis elegans by sequence and cellular distribution. In tether function assays, 4E-T represses bound mRNA translation, in a manner independent of these localization sequences, or of endogenous P-bodies. Quantitative polymerase chain reaction and northern blot analysis verified that bound mRNA remained intact and polyadenylated. Ectopic 4E-T reduces translation globally in a manner dependent on eIF4E binding its consensus Y30X4L site. In contrast, tethered 4E-T continued to repress translation when eIF4E-binding was prevented by mutagenesis of YX4L, and modestly enhanced the decay of bound mRNA, compared with wild-type 4E-T, mediated by increased binding of CNOT1/7 deadenylase subunits. As depleting 4E-T from HeLa cells increased steady-state translation, in part due to relief of microRNA-mediated silencing, this work demonstrates the conserved yet unconventional mechanism of 4E-T silencing of particular subsets of mRNAs.
Figures
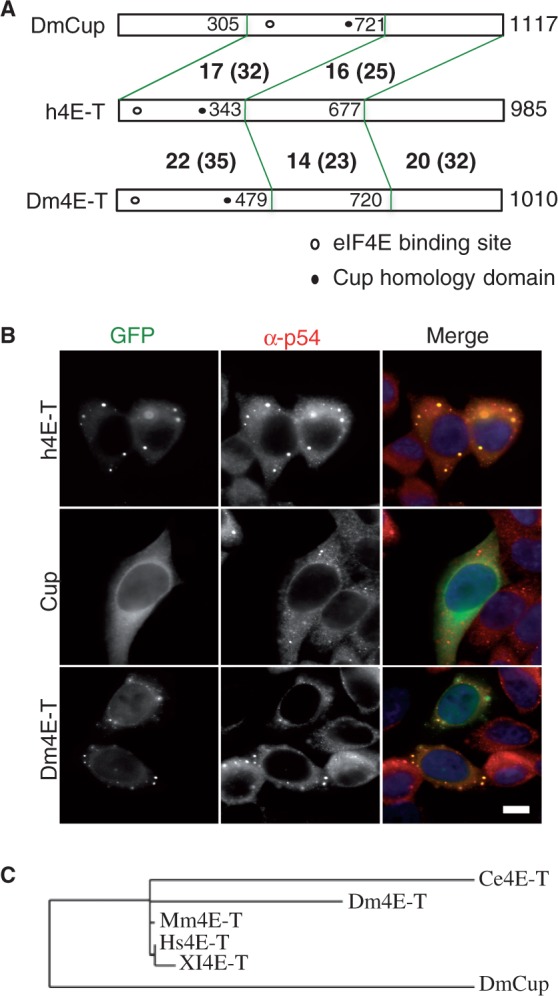
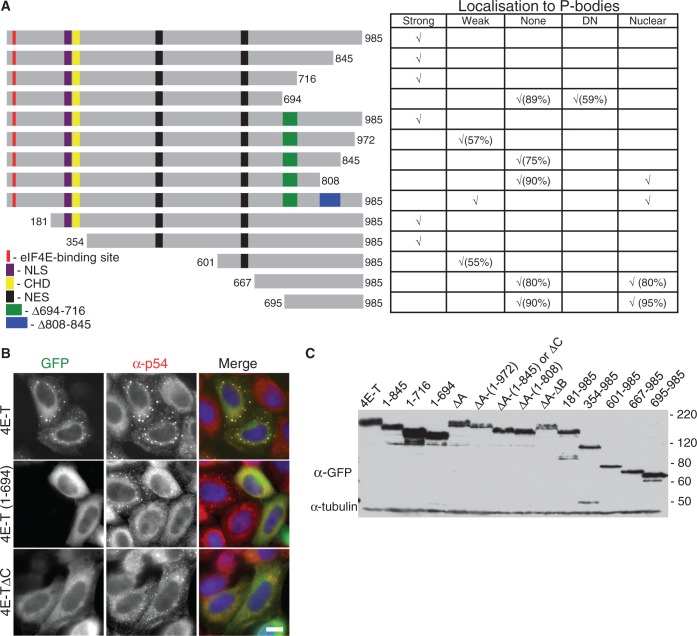
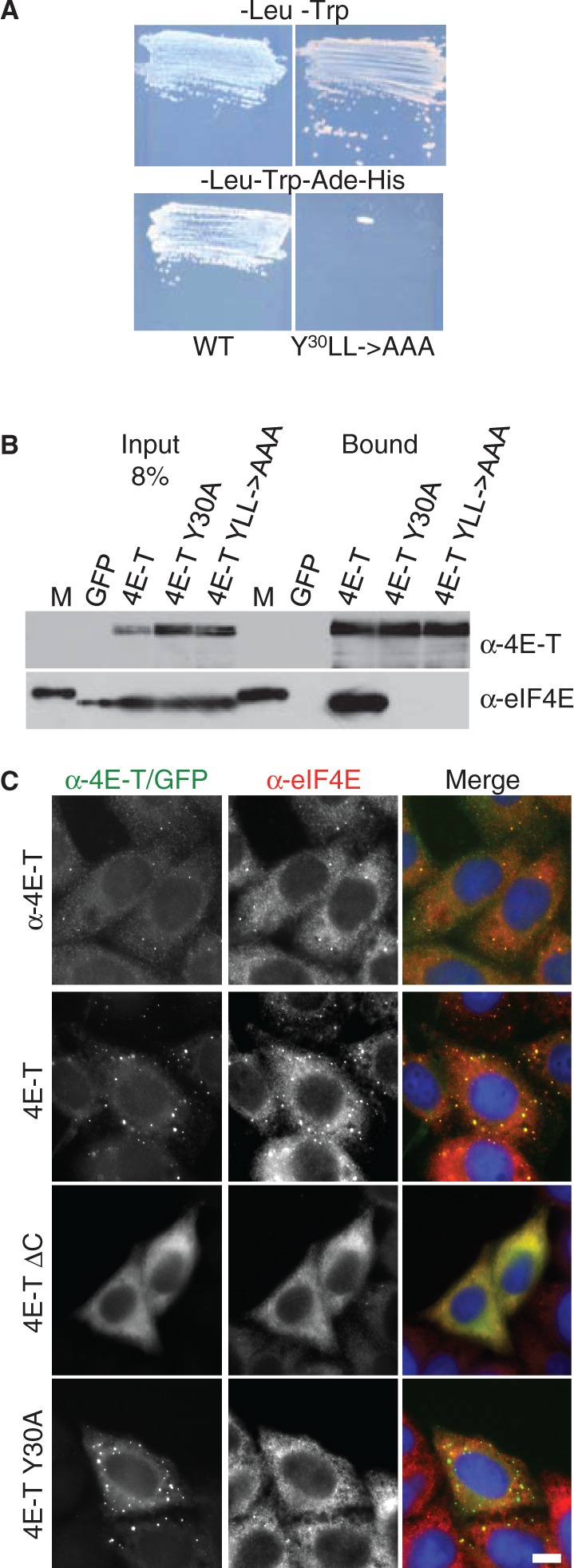
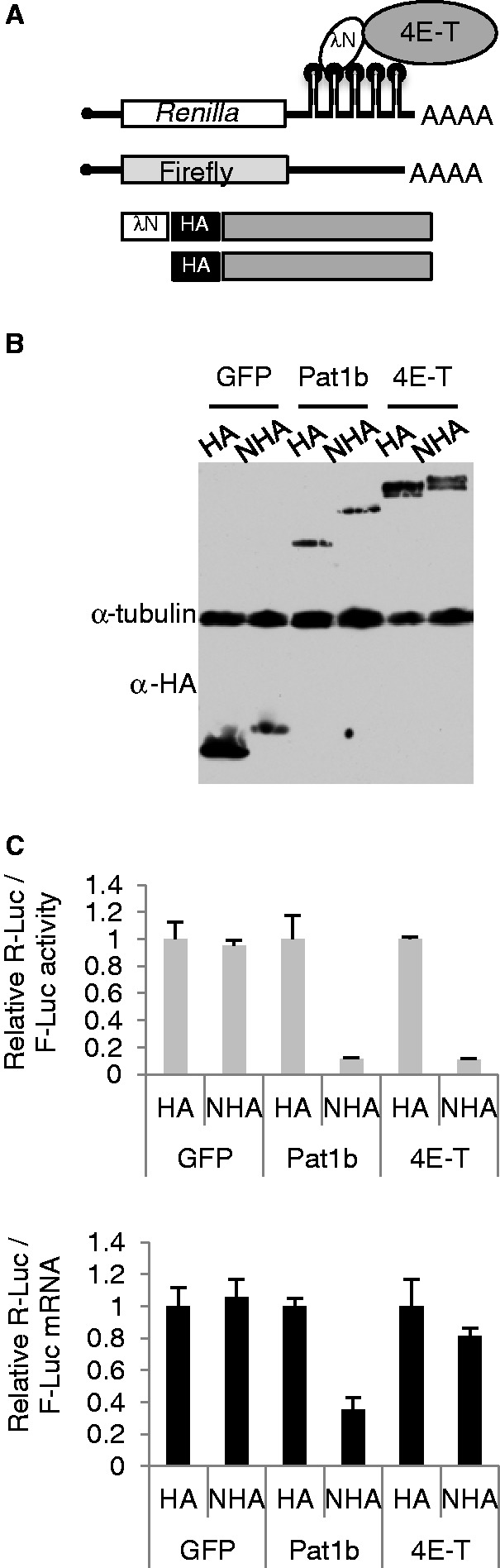

Similar articles
-
The DDX6-4E-T interaction mediates translational repression and P-body assembly.Nucleic Acids Res. 2016 Jul 27;44(13):6318-34. doi: 10.1093/nar/gkw565. Epub 2016 Jun 24. Nucleic Acids Res. 2016. PMID: 27342281 Free PMC article.
-
A role for the eIF4E-binding protein 4E-T in P-body formation and mRNA decay.J Cell Biol. 2005 Sep 12;170(6):913-24. doi: 10.1083/jcb.200504039. J Cell Biol. 2005. PMID: 16157702 Free PMC article.
-
Ectopic expression of eIF4E-transporter triggers the movement of eIF4E into P-bodies, inhibiting steady-state translation but not the pioneer round of translation.Biochem Biophys Res Commun. 2008 May 16;369(4):1160-5. doi: 10.1016/j.bbrc.2008.03.017. Epub 2008 Mar 14. Biochem Biophys Res Commun. 2008. PMID: 18343217
-
Does phosphorylation of the cap-binding protein eIF4E play a role in translation initiation?Eur J Biochem. 2002 Nov;269(22):5350-9. doi: 10.1046/j.1432-1033.2002.03291.x. Eur J Biochem. 2002. PMID: 12423333 Free PMC article. Review.
-
eIF4E: new family members, new binding partners, new roles.J Biol Chem. 2009 Jun 19;284(25):16711-16715. doi: 10.1074/jbc.R900002200. Epub 2009 Feb 23. J Biol Chem. 2009. PMID: 19237539 Free PMC article. Review.
Cited by
-
Roles of the CCR4-Not complex in translation and dynamics of co-translation events.Wiley Interdiscip Rev RNA. 2023 Nov 27;15(1):e1827. doi: 10.1002/wrna.1827. Online ahead of print. Wiley Interdiscip Rev RNA. 2023. PMID: 38009591 Free PMC article. Review.
-
Mechanistic Insights into MicroRNA-Mediated Gene Silencing.Cold Spring Harb Perspect Biol. 2019 Mar 1;11(3):a032771. doi: 10.1101/cshperspect.a032771. Cold Spring Harb Perspect Biol. 2019. PMID: 29959194 Free PMC article. Review.
-
dCas13-mediated translational repression for accurate gene silencing in mammalian cells.Nat Commun. 2024 Mar 11;15(1):2205. doi: 10.1038/s41467-024-46412-7. Nat Commun. 2024. PMID: 38467613 Free PMC article.
-
Extracellular MicroRNAs as Intercellular Mediators and Noninvasive Biomarkers of Cancer.Cancers (Basel). 2020 Nov 20;12(11):3455. doi: 10.3390/cancers12113455. Cancers (Basel). 2020. PMID: 33233600 Free PMC article. Review.
-
Using the Tether Function Assay to Identify Potential Regulators of mRNA Translation and mRNA Decay.Methods Mol Biol. 2025;2863:265-280. doi: 10.1007/978-1-0716-4176-7_16. Methods Mol Biol. 2025. PMID: 39535715
References
-
- Minshall N, Reiter M-H, Weil D, Standart N. CPEB interacts with an ovary-specific eIF4E and 4E-T in early Xenopus oocytes. J. Biol. Chem. 2007;282:37389–37401. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MC_UP_A600_1024/MRC_/Medical Research Council/United Kingdom
- BB/J00779X/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 084885/Z/08/Z/WT_/Wellcome Trust/United Kingdom
- G0902052/MRC_/Medical Research Council/United Kingdom
- MC_EX_G0902052/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
