

Loss of col8a1a function during zebrafish embryogenesis results in congenital vertebral malformations
- PMID: 24333517
- PMCID: PMC3938106
- DOI: 10.1016/j.ydbio.2013.11.028
Loss of col8a1a function during zebrafish embryogenesis results in congenital vertebral malformations
Abstract
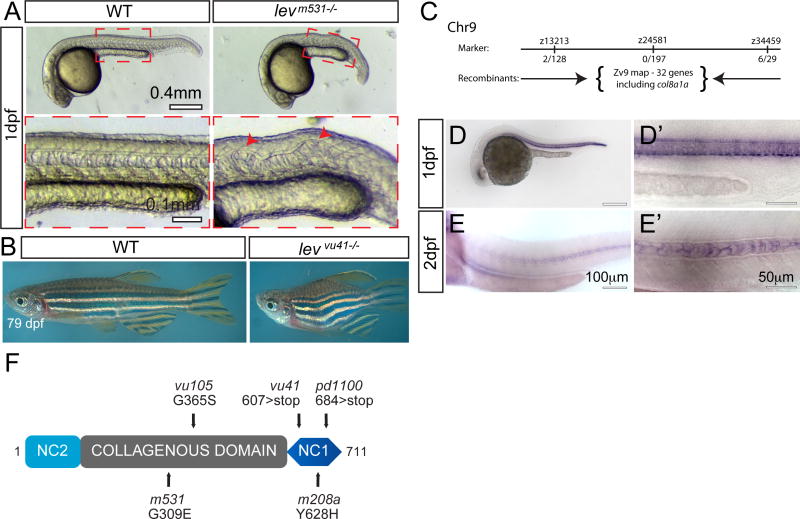
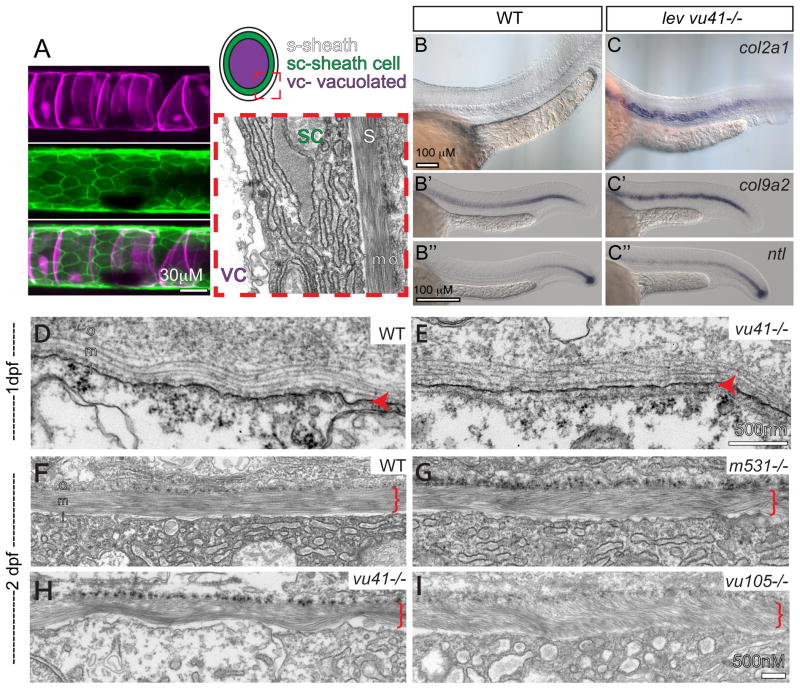
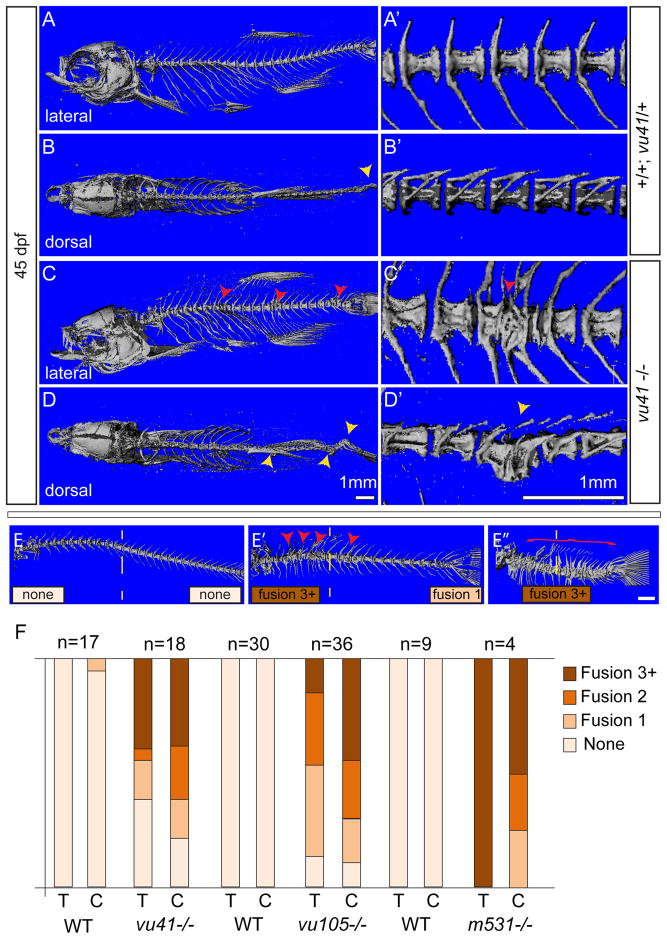
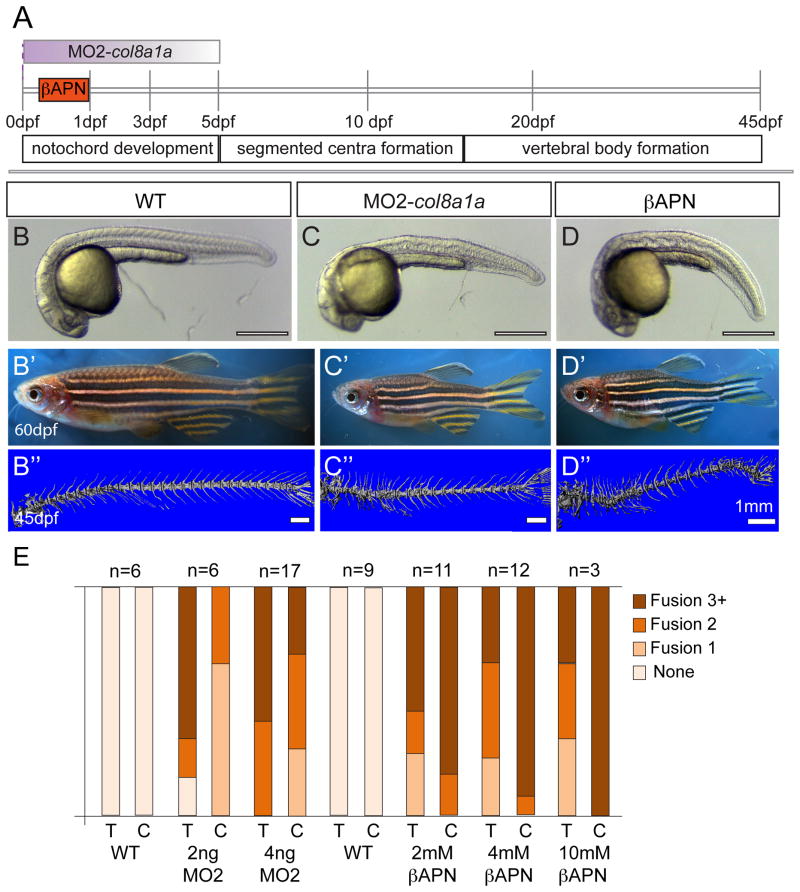
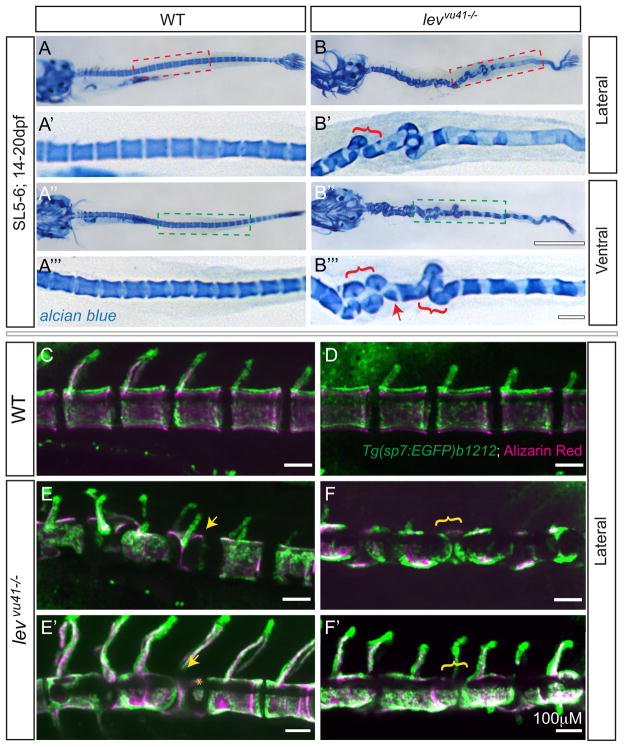
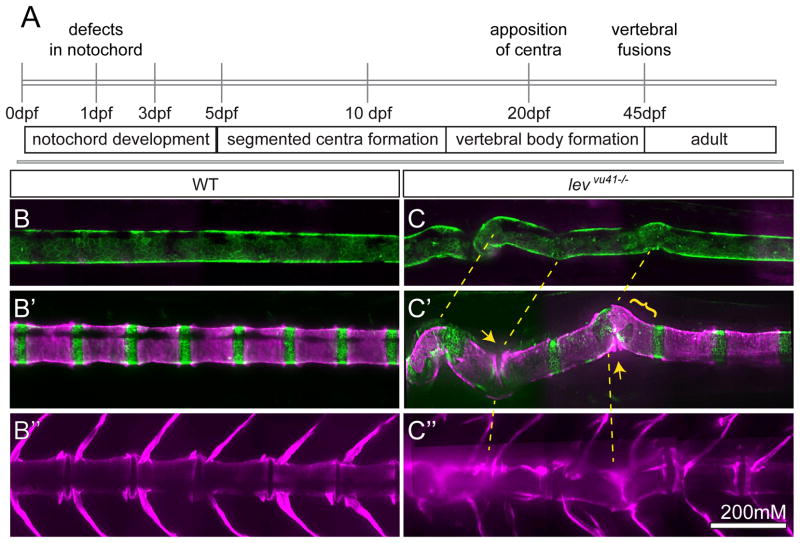
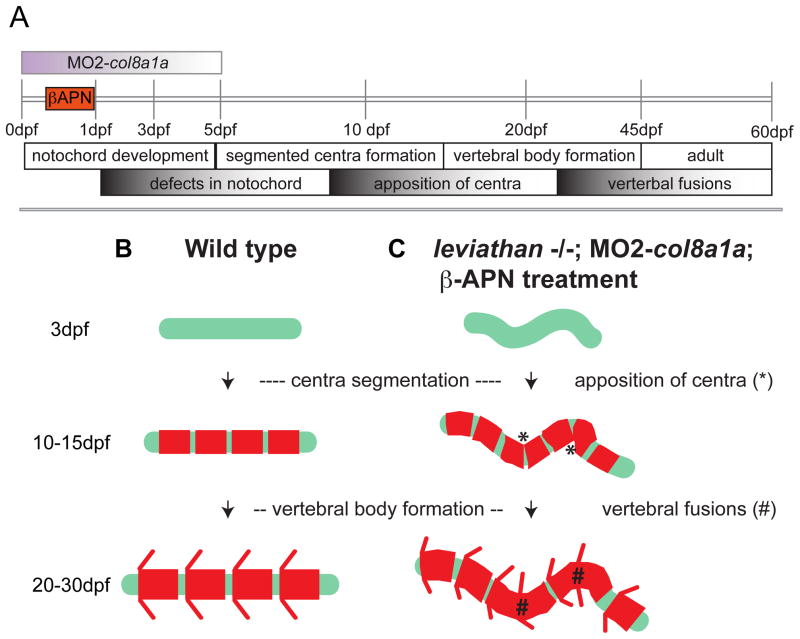
Congenital vertebral malformations (CVM) occur in 1 in 1000 live births and in many cases can cause spinal deformities, such as scoliosis, and result in disability and distress of affected individuals. Many severe forms of the disease, such as spondylocostal dystostosis, are recessive monogenic traits affecting somitogenesis, however the etiologies of the majority of CVM cases remain undetermined. Here we demonstrate that morphological defects of the notochord in zebrafish can generate congenital-type spine defects. We characterize three recessive zebrafish leviathan/col8a1a mutant alleles ((m531, vu41, vu105)) that disrupt collagen type VIII alpha1a (col8a1a), and cause folding of the embryonic notochord and consequently adult vertebral column malformations. Furthermore, we provide evidence that a transient loss of col8a1a function or inhibition of Lysyl oxidases with drugs during embryogenesis was sufficient to generate vertebral fusions and scoliosis in the adult spine. Using periodic imaging of individual zebrafish, we correlate focal notochord defects of the embryo with vertebral malformations (VM) in the adult. Finally, we show that bends and kinks in the notochord can lead to aberrant apposition of osteoblasts normally confined to well-segmented areas of the developing vertebral bodies. Our results afford a novel mechanism for the formation of VM, independent of defects of somitogenesis, resulting from aberrant bone deposition at regions of misshapen notochord tissue.
Keywords: Collagen; Notochord; Osteoblast; Vertebral malformations.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Essential role for the alpha 1 chain of type VIII collagen in zebrafish notochord formation.Dev Dyn. 2008 Dec;237(12):3715-26. doi: 10.1002/dvdy.21779. Dev Dyn. 2008. PMID: 19035365 Free PMC article.
-
Notochord vacuoles absorb compressive bone growth during zebrafish spine formation.Elife. 2020 Jan 29;9:e51221. doi: 10.7554/eLife.51221. Elife. 2020. PMID: 31995030 Free PMC article.
-
Dstyk mutation leads to congenital scoliosis-like vertebral malformations in zebrafish via dysregulated mTORC1/TFEB pathway.Nat Commun. 2020 Jan 24;11(1):479. doi: 10.1038/s41467-019-14169-z. Nat Commun. 2020. PMID: 31980602 Free PMC article.
-
Progress and perspective of TBX6 gene in congenital vertebral malformations.Oncotarget. 2016 Aug 30;7(35):57430-57441. doi: 10.18632/oncotarget.10619. Oncotarget. 2016. PMID: 27437870 Free PMC article. Review.
-
Disordered vertebral and rib morphology in pudgy mice. Structural relationships to human scoliosis.Adv Anat Embryol Cell Biol. 2016;221:1-123. doi: 10.1007/978-3-319-43151-2_1. Adv Anat Embryol Cell Biol. 2016. PMID: 27655002 Review.
Cited by
-
Zebrafish: an important model for understanding scoliosis.Cell Mol Life Sci. 2022 Sep 4;79(9):506. doi: 10.1007/s00018-022-04534-5. Cell Mol Life Sci. 2022. PMID: 36059018 Free PMC article. Review.
-
Development of a straight vertebrate body axis.Development. 2020 Oct 6;147(21):dev175794. doi: 10.1242/dev.175794. Development. 2020. PMID: 33023886 Free PMC article. Review.
-
Spinal scoliosis: insights into developmental mechanisms and animal models.Spine Deform. 2025 Jan;13(1):7-18. doi: 10.1007/s43390-024-00941-9. Epub 2024 Aug 20. Spine Deform. 2025. PMID: 39164474 Free PMC article. Review.
-
Stat3/Cdc25a-dependent cell proliferation promotes embryonic axis extension during zebrafish gastrulation.PLoS Genet. 2017 Feb 21;13(2):e1006564. doi: 10.1371/journal.pgen.1006564. eCollection 2017 Feb. PLoS Genet. 2017. PMID: 28222105 Free PMC article.
-
Unraveling the genetic architecture of congenital vertebral malformation with reference to the developing spine.Nat Commun. 2024 Feb 6;15(1):1125. doi: 10.1038/s41467-024-45442-5. Nat Commun. 2024. PMID: 38321032 Free PMC article.
References
-
- Adams DS, Keller R, Koehl MA. The mechanics of notochord elongation, straightening and stiffening in the embryo of Xenopus laevis. Development. 1990;110:115–30. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
