

TGF-β signalling is required for CD4⁺ T cell homeostasis but dispensable for regulatory T cell function
- PMID: 24115907
- PMCID: PMC3792861
- DOI: 10.1371/journal.pbio.1001674
TGF-β signalling is required for CD4⁺ T cell homeostasis but dispensable for regulatory T cell function
Abstract
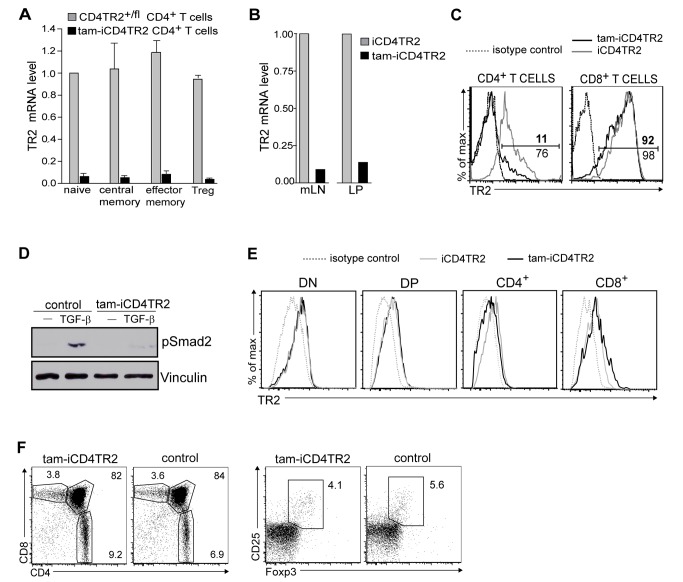
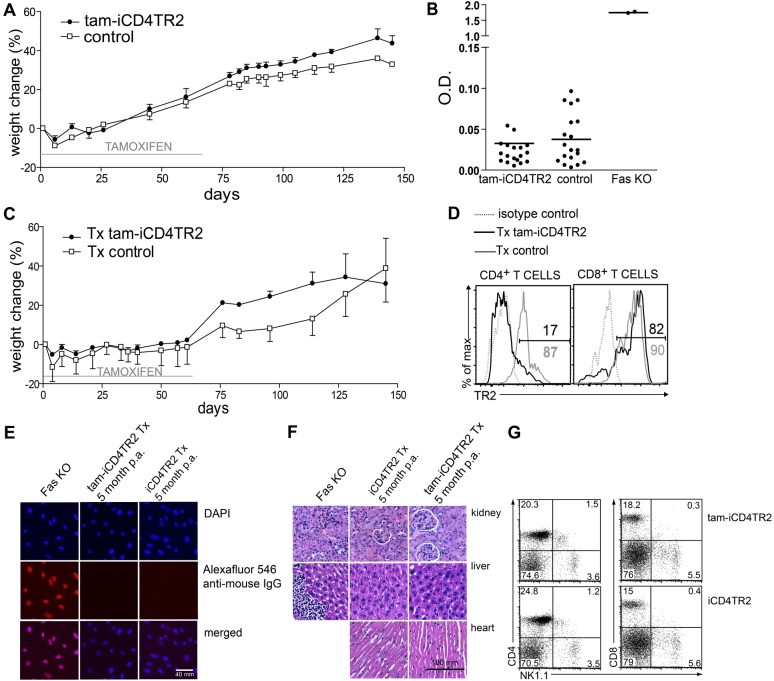
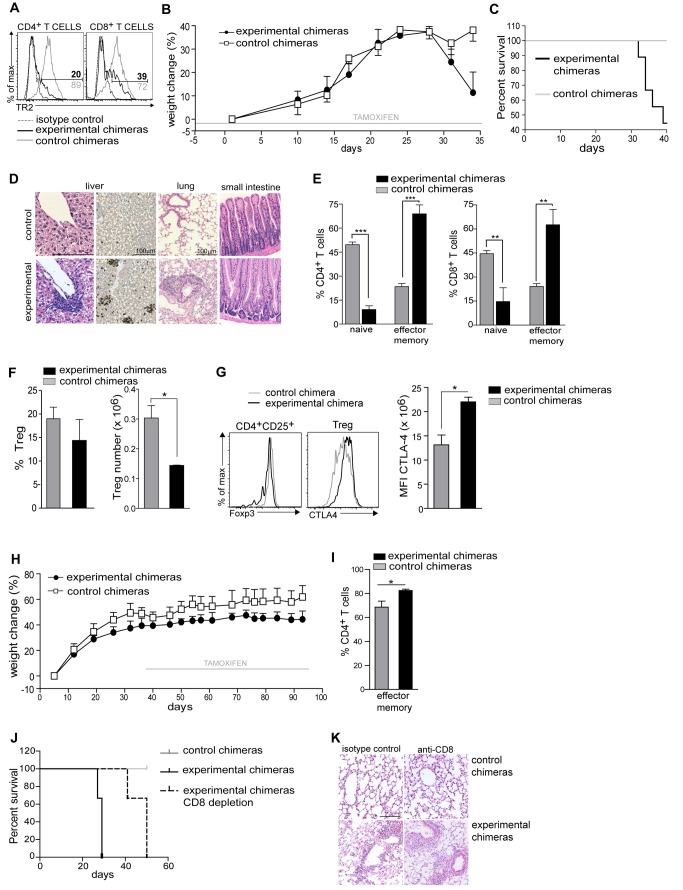
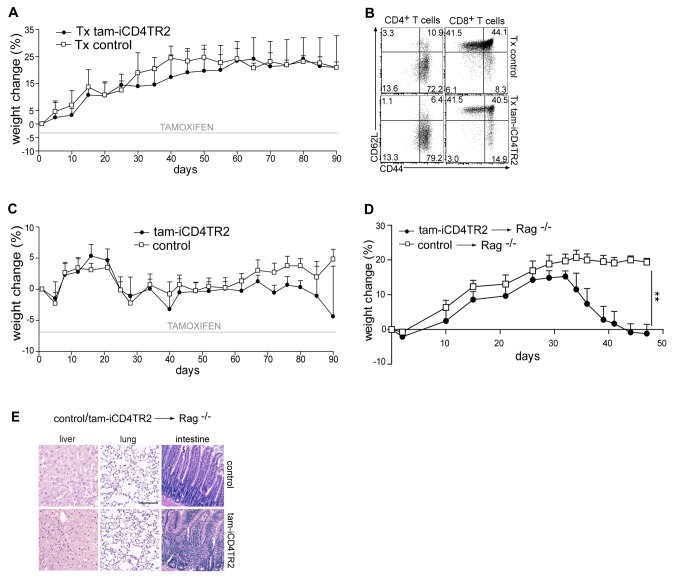
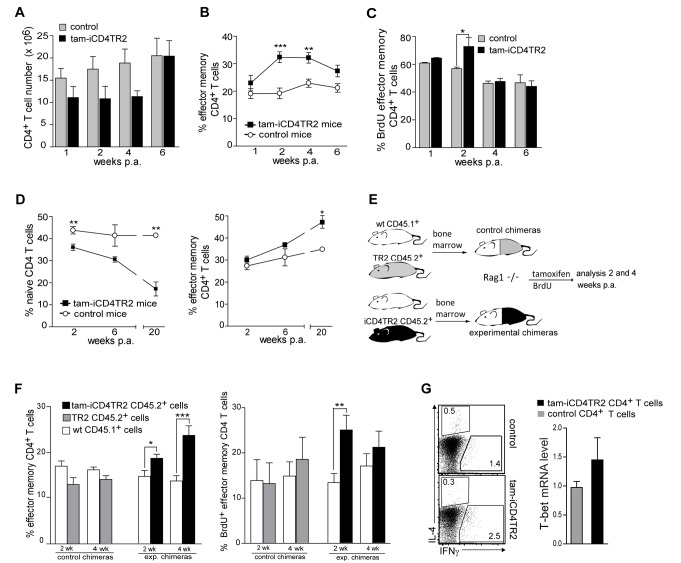
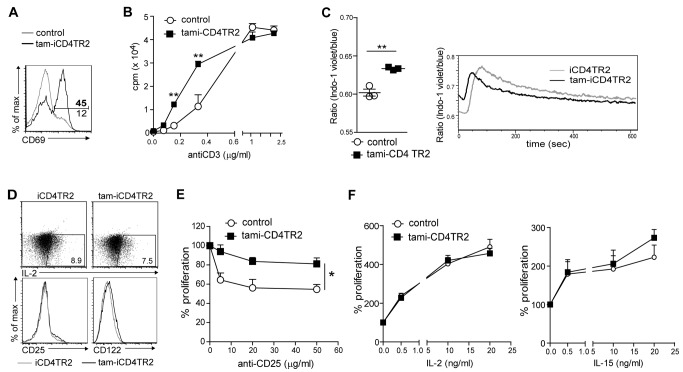
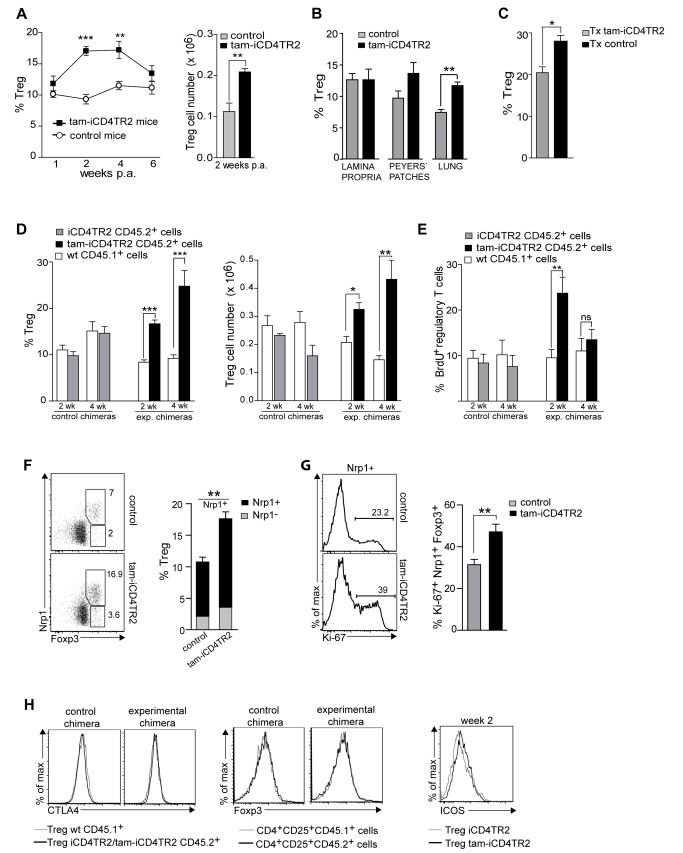
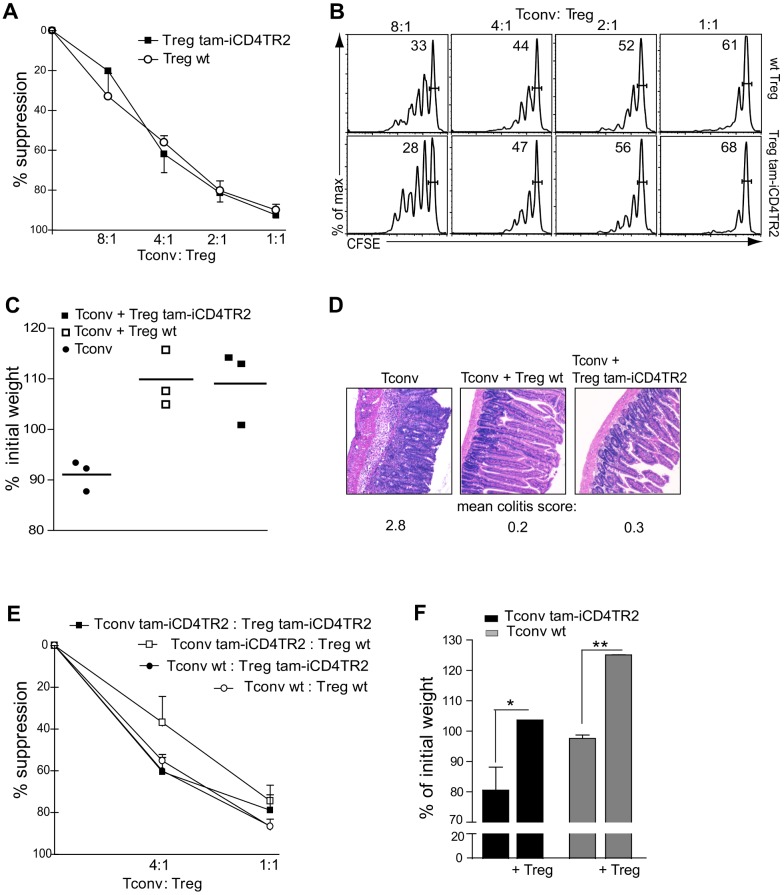
TGF-β is widely held to be critical for the maintenance and function of regulatory T (T(reg)) cells and thus peripheral tolerance. This is highlighted by constitutive ablation of TGF-β receptor (TR) during thymic development in mice, which leads to a lethal autoimmune syndrome. Here we describe that TGF-β-driven peripheral tolerance is not regulated by TGF-β signalling on mature CD4⁺ T cells. Inducible TR2 ablation specifically on CD4⁺ T cells did not result in a lethal autoinflammation. Transfer of these TR2-deficient CD4⁺ T cells to lymphopenic recipients resulted in colitis, but not overt autoimmunity. In contrast, thymic ablation of TR2 in combination with lymphopenia led to lethal multi-organ inflammation. Interestingly, deletion of TR2 on mature CD4⁺ T cells does not result in the collapse of the T(reg) cell population as observed in constitutive models. Instead, a pronounced enlargement of both regulatory and effector memory T cell pools was observed. This expansion is cell-intrinsic and seems to be caused by increased T cell receptor sensitivity independently of common gamma chain-dependent cytokine signals. The expression of Foxp3 and other regulatory T cells markers was not dependent on TGF-β signalling and the TR2-deficient T(reg) cells retained their suppressive function both in vitro and in vivo. In summary, absence of TGF-β signalling on mature CD4⁺ T cells is not responsible for breakdown of peripheral tolerance, but rather controls homeostasis of mature T cells in adult mice.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
TGF-β signaling to T cells inhibits autoimmunity during lymphopenia-driven proliferation.Nat Immunol. 2012 May 27;13(7):667-73. doi: 10.1038/ni.2319. Nat Immunol. 2012. PMID: 22634866 Free PMC article.
-
Excessive Th1 responses due to the absence of TGF-β signaling cause autoimmune diabetes and dysregulated Treg cell homeostasis.Proc Natl Acad Sci U S A. 2013 Apr 23;110(17):6961-6. doi: 10.1073/pnas.1304498110. Epub 2013 Apr 8. Proc Natl Acad Sci U S A. 2013. PMID: 23569233 Free PMC article.
-
TGF-beta1 maintains suppressor function and Foxp3 expression in CD4+CD25+ regulatory T cells.J Exp Med. 2005 Apr 4;201(7):1061-7. doi: 10.1084/jem.20042276. J Exp Med. 2005. PMID: 15809351 Free PMC article.
-
Regulatory T-cells: diverse phenotypes integral to immune homeostasis and suppression.Toxicol Pathol. 2012;40(2):186-204. doi: 10.1177/0192623311430693. Epub 2012 Jan 5. Toxicol Pathol. 2012. PMID: 22222887 Review.
-
Transforming growth factor-beta: an important role in CD4+CD25+ regulatory T cells and immune tolerance.Autoimmunity. 2006 Jun;39(4):269-76. doi: 10.1080/08916930600753903. Autoimmunity. 2006. PMID: 16891215 Review.
Cited by
-
TGFβ control of immune responses in cancer: a holistic immuno-oncology perspective.Nat Rev Immunol. 2023 Jun;23(6):346-362. doi: 10.1038/s41577-022-00796-z. Epub 2022 Nov 15. Nat Rev Immunol. 2023. PMID: 36380023 Free PMC article. Review.
-
Role of regulatory T cell in the pathogenesis of inflammatory bowel disease.World J Gastroenterol. 2016 Feb 21;22(7):2195-205. doi: 10.3748/wjg.v22.i7.2195. World J Gastroenterol. 2016. PMID: 26900284 Free PMC article. Review.
-
Indoleamine 2,3-Dioxygenase 1 Inhibitor-Loaded Nanosheets Enhance CAR-T Cell Function in Esophageal Squamous Cell Carcinoma.Front Immunol. 2021 Mar 22;12:661357. doi: 10.3389/fimmu.2021.661357. eCollection 2021. Front Immunol. 2021. PMID: 33828565 Free PMC article.
-
Prior to Peripheral Tolerance, Newly Generated CD4 T Cells Maintain Dangerous Autoimmune Potential: Fas- and Perforin-Independent Autoimmunity Controlled by Programmed Death-1.Front Immunol. 2018 Jan 24;9:12. doi: 10.3389/fimmu.2018.00012. eCollection 2018. Front Immunol. 2018. PMID: 29416537 Free PMC article.
-
TGF-β receptor maintains CD4 T helper cell identity during chronic viral infections.J Clin Invest. 2016 Oct 3;126(10):3799-3813. doi: 10.1172/JCI87041. Epub 2016 Sep 6. J Clin Invest. 2016. PMID: 27599295 Free PMC article.
References
-
- Oshima M, Oshima H, Taketo MM (1996) TGF-beta receptor type II deficiency results in defects of yolk sac hematopoiesis and vasculogenesis. Dev Biol 179: 297–302. - PubMed
-
- Gorelik L, Flavell RA (2000) Abrogation of TGFbeta signaling in T cells leads to spontaneous T cell differentiation and autoimmune disease. Immunity 12: 171–181. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
