

Neutrophils express oncomodulin and promote optic nerve regeneration
- PMID: 24027282
- PMCID: PMC3771038
- DOI: 10.1523/JNEUROSCI.5511-12.2013
Neutrophils express oncomodulin and promote optic nerve regeneration
Abstract
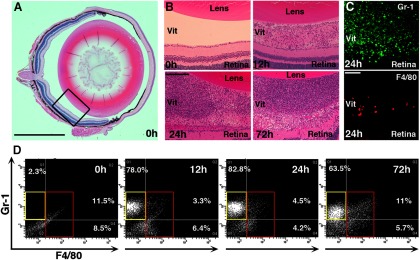
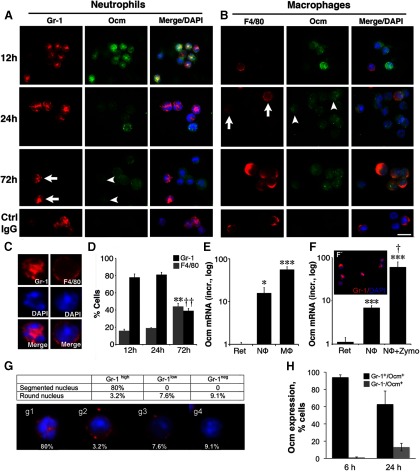
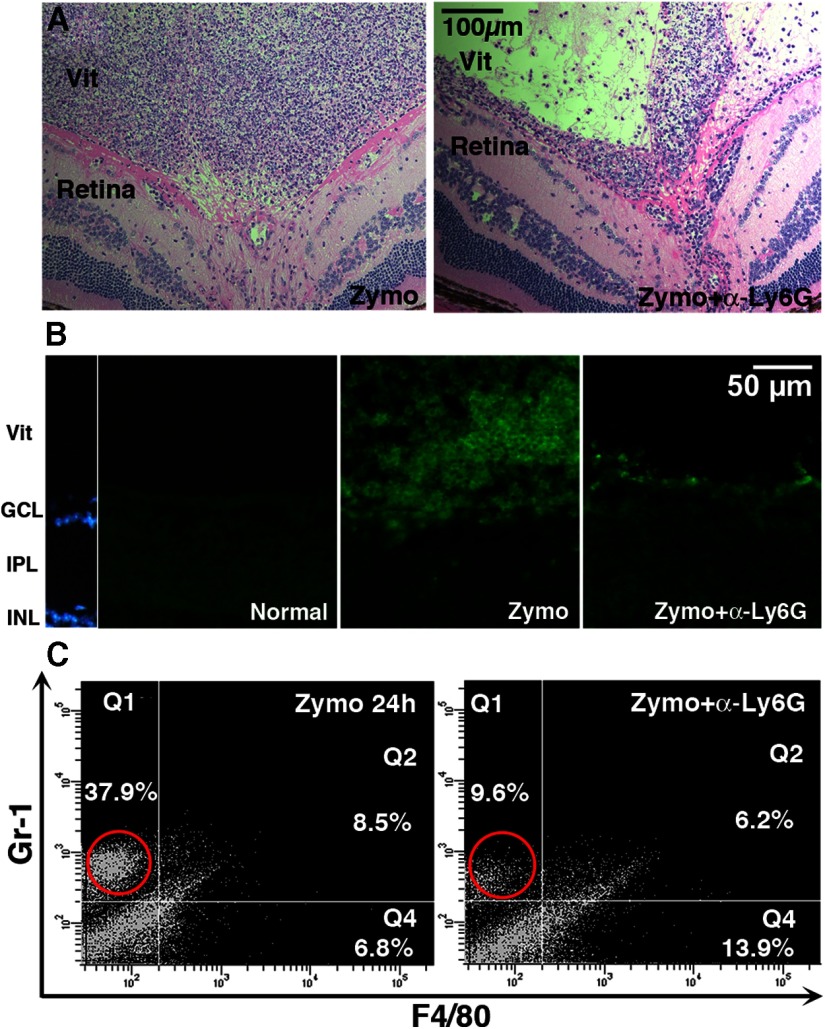
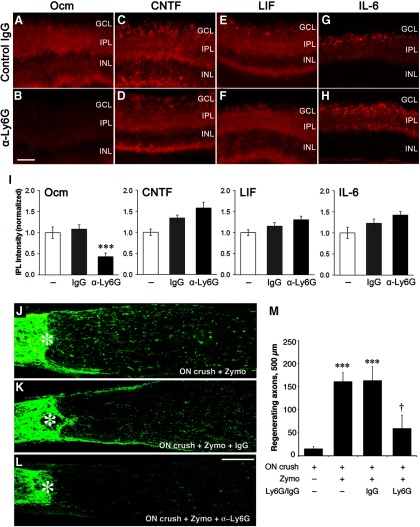
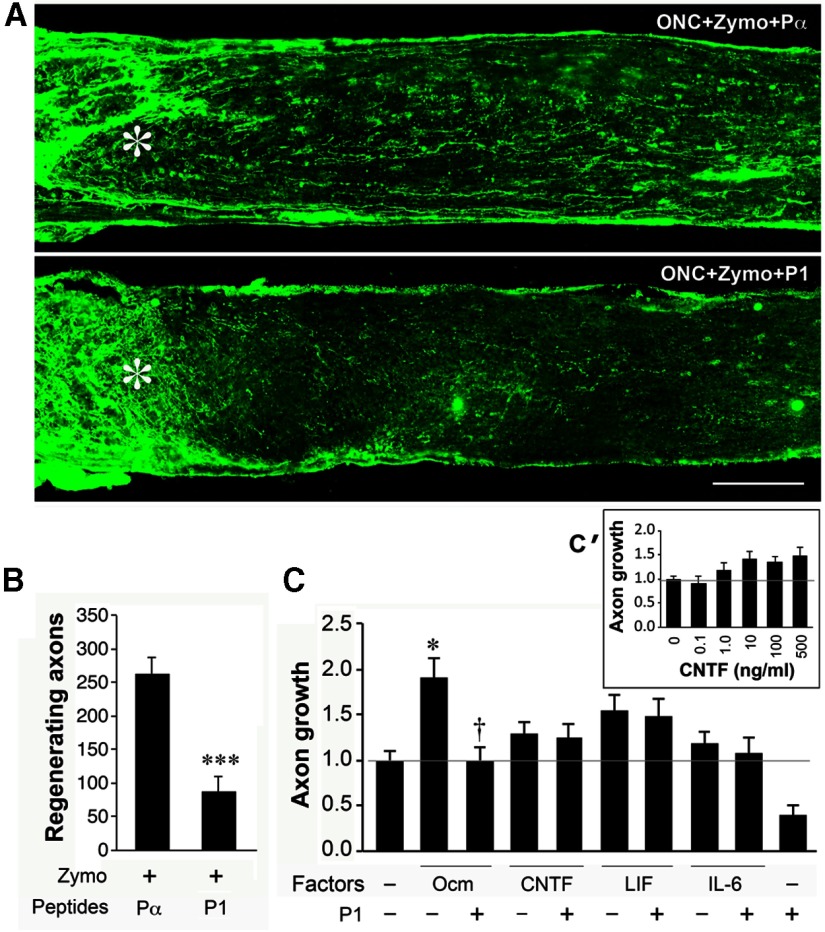
Although neurons are normally unable to regenerate their axons after injury to the CNS, this situation can be partially reversed by activating the innate immune system. In a widely studied instance of this phenomenon, proinflammatory agents have been shown to cause retinal ganglion cells, the projection neurons of the eye, to regenerate lengthy axons through the injured optic nerve. However, the role of different molecules and cell populations in mediating this phenomenon remains unclear. We show here that neutrophils, the first responders of the innate immune system, play a central role in inflammation-induced regeneration. Numerous neutrophils enter the mouse eye within a few hours of inducing an inflammatory reaction and express high levels of the atypical growth factor oncomodulin (Ocm). Immunodepletion of neutrophils diminished Ocm levels in the eye without altering levels of CNTF, leukemia inhibitory factor, or IL-6, and suppressed the proregenerative effects of inflammation. A peptide antagonist of Ocm suppressed regeneration as effectively as neutrophil depletion. Macrophages enter the eye later in the inflammatory process but appear to be insufficient to stimulate extensive regeneration in the absence of neutrophils. These data provide the first evidence that neutrophils are a major source of Ocm and can promote axon regeneration in the CNS.
Figures
Comment in
-
Neutrophil contribution in facilitating optic nerve regeneration.J Neurosci. 2014 Jan 22;34(4):1081-2. doi: 10.1523/JNEUROSCI.4827-13.2014. J Neurosci. 2014. PMID: 24453300 Free PMC article. No abstract available.
Similar articles
-
Neuroprotective and axon growth-promoting effects following inflammatory stimulation on mature retinal ganglion cells in mice depend on ciliary neurotrophic factor and leukemia inhibitory factor.J Neurosci. 2009 Nov 11;29(45):14334-41. doi: 10.1523/JNEUROSCI.2770-09.2009. J Neurosci. 2009. PMID: 19906980 Free PMC article.
-
Oncomodulin is a macrophage-derived signal for axon regeneration in retinal ganglion cells.Nat Neurosci. 2006 Jun;9(6):843-52. doi: 10.1038/nn1701. Epub 2006 May 14. Nat Neurosci. 2006. PMID: 16699509
-
Oncomodulin links inflammation to optic nerve regeneration.Proc Natl Acad Sci U S A. 2009 Nov 17;106(46):19587-92. doi: 10.1073/pnas.0907085106. Epub 2009 Oct 29. Proc Natl Acad Sci U S A. 2009. PMID: 19875691 Free PMC article.
-
Rewiring the injured CNS: lessons from the optic nerve.Exp Neurol. 2008 Feb;209(2):389-98. doi: 10.1016/j.expneurol.2007.05.025. Epub 2007 Jun 7. Exp Neurol. 2008. PMID: 17610877 Free PMC article. Review.
-
Reaching the brain: Advances in optic nerve regeneration.Exp Neurol. 2017 Jan;287(Pt 3):365-373. doi: 10.1016/j.expneurol.2015.12.015. Epub 2015 Dec 31. Exp Neurol. 2017. PMID: 26746987 Review.
Cited by
-
Early retinal inflammatory biomarkers in the middle cerebral artery occlusion model of ischemic stroke.Mol Vis. 2016 Jun 4;22:575-88. eCollection 2016. Mol Vis. 2016. PMID: 27293375 Free PMC article.
-
Beyond host defense and tissue injury: the emerging role of neutrophils in tissue repair.Am J Physiol Cell Physiol. 2024 Mar 1;326(3):C661-C683. doi: 10.1152/ajpcell.00652.2023. Epub 2024 Jan 8. Am J Physiol Cell Physiol. 2024. PMID: 38189129 Free PMC article. Review.
-
Neutrophils in tissue injury and repair.Cell Tissue Res. 2018 Mar;371(3):531-539. doi: 10.1007/s00441-017-2785-7. Epub 2018 Jan 30. Cell Tissue Res. 2018. PMID: 29383445 Free PMC article. Review.
-
Immunomodulation by the combination of statin and matrix-bound nanovesicle enhances optic nerve regeneration.NPJ Regen Med. 2024 Oct 26;9(1):31. doi: 10.1038/s41536-024-00374-y. NPJ Regen Med. 2024. PMID: 39461953 Free PMC article.
-
Neutrophils in Ocular Diseases.Int J Mol Sci. 2024 Jul 15;25(14):7736. doi: 10.3390/ijms25147736. Int J Mol Sci. 2024. PMID: 39062975 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical