

The Nlrp3 inflammasome regulates acute graft-versus-host disease
- PMID: 23980097
- PMCID: PMC3782050
- DOI: 10.1084/jem.20130084
The Nlrp3 inflammasome regulates acute graft-versus-host disease
Abstract
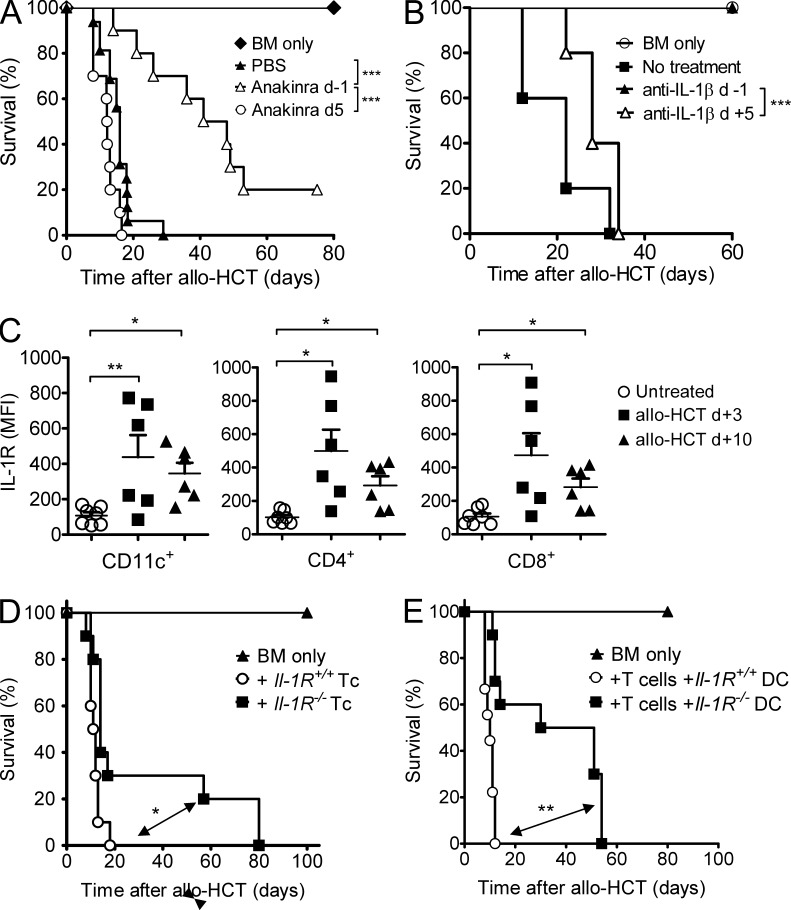
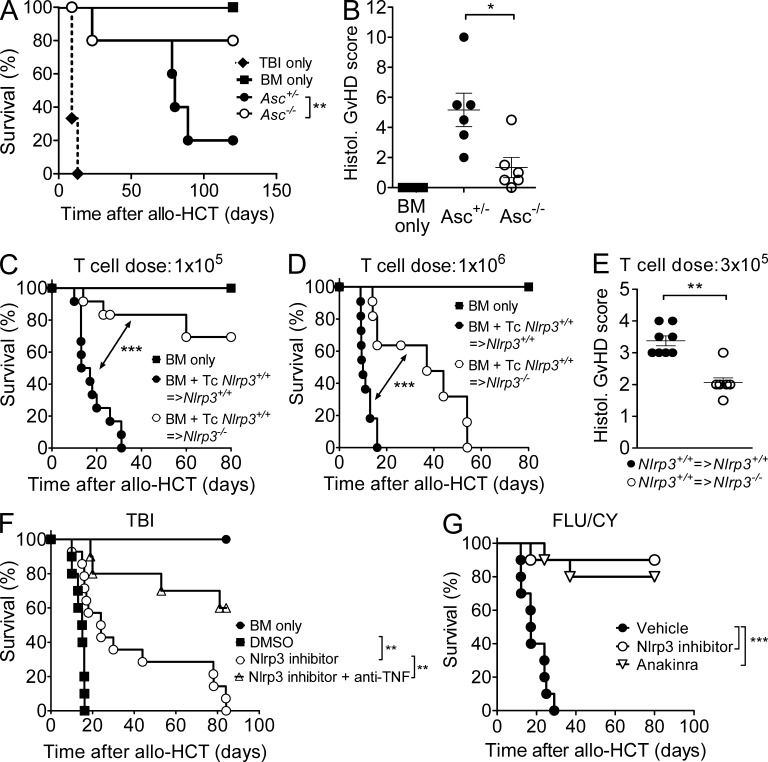
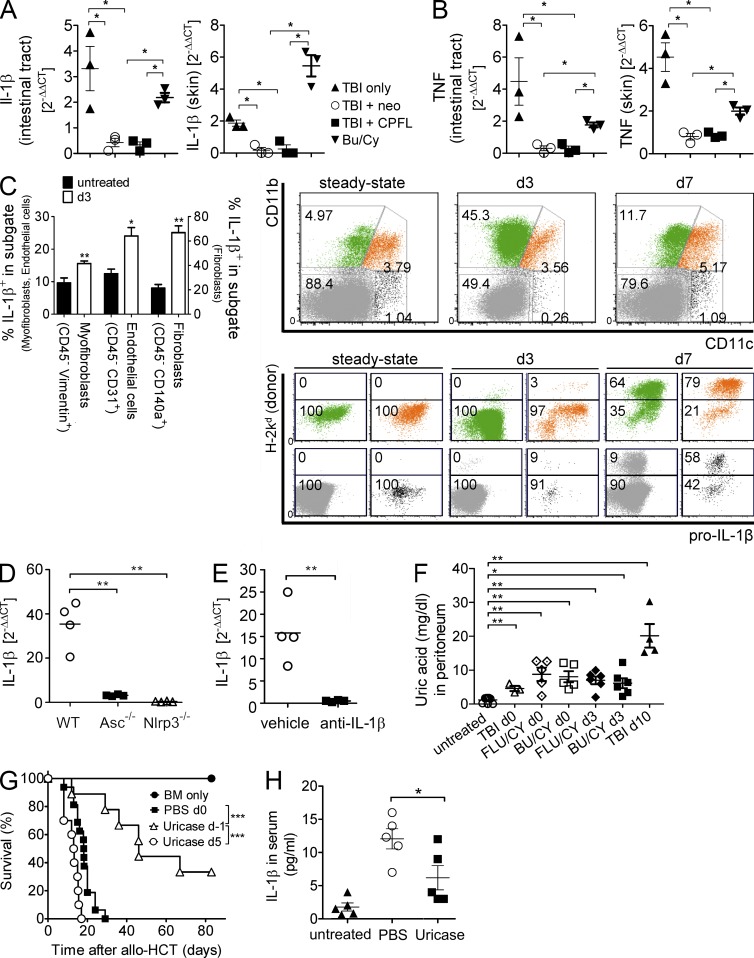
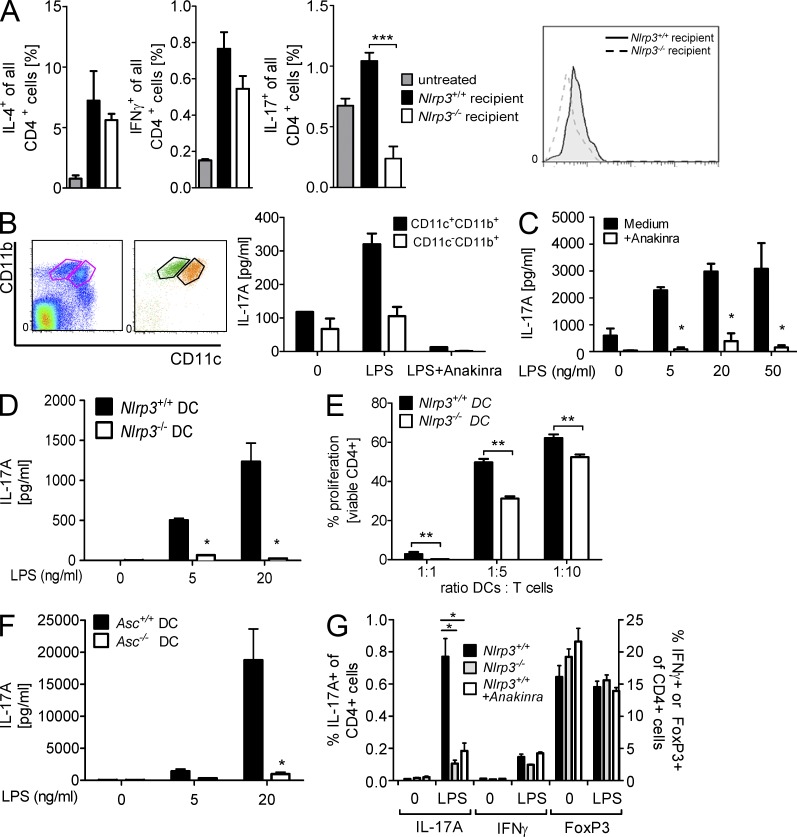
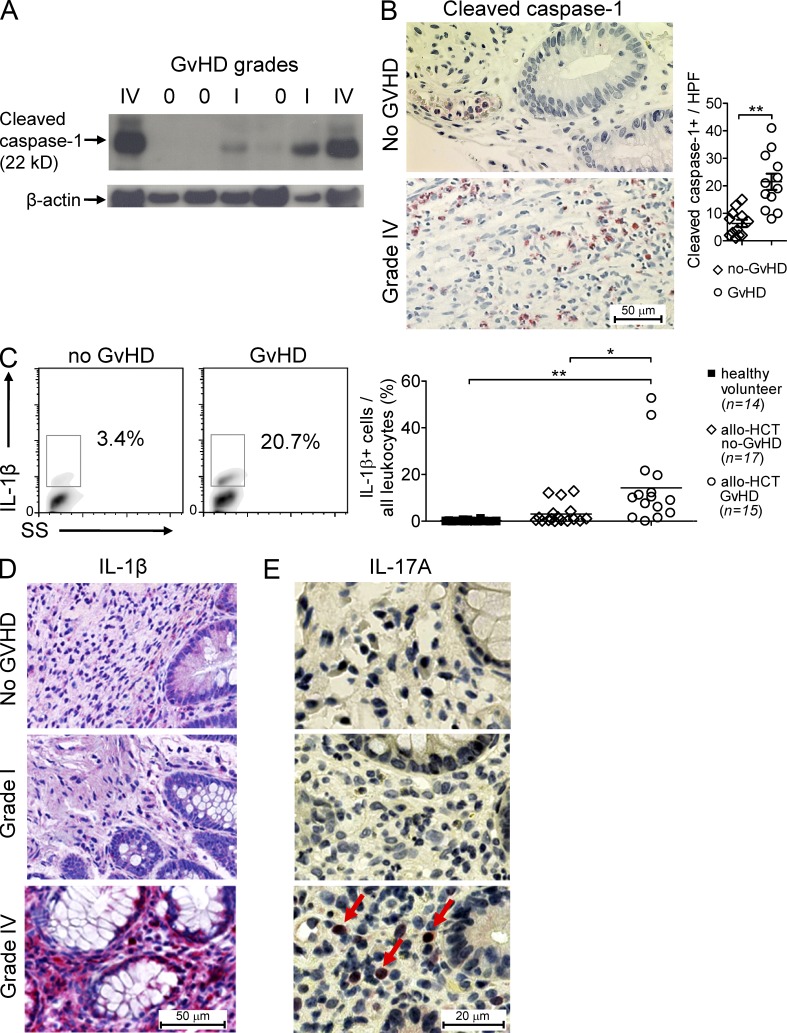
The success of allogeneic hematopoietic cell transplantation is limited by acute graft-versus-host disease (GvHD), a severe complication accompanied by high mortality rates. Yet, the molecular mechanisms initiating this disease remain poorly defined. In this study, we show that, after conditioning therapy, intestinal commensal bacteria and the damage-associated molecular pattern uric acid contribute to Nlrp3 inflammasome-mediated IL-1β production and that gastrointestinal decontamination and uric acid depletion reduced GvHD severity. Early blockade of IL-1β or genetic deficiency of the IL-1 receptor in dendritic cells (DCs) and T cells improved survival. The Nlrp3 inflammasome components Nlrp3 and Asc, which are required for pro-IL-1β cleavage, were critical for the full manifestation of GvHD. In transplanted mice, IL-1β originated from multiple intestinal cell compartments and exerted its effects on DCs and T cells, the latter being preferentially skewed toward Th17. Compatible with these mouse data, increased levels of active caspase-1 and IL-1β were found in circulating leukocytes and intestinal GvHD lesions of patients. Thus, the identification of a crucial role for the Nlrp3 inflammasome sheds new light on the pathogenesis of GvHD and opens a potential new avenue for the targeted therapy of this severe complication.
Figures
Similar articles
-
Activation of the NLRP3 inflammasome by group B streptococci.J Immunol. 2012 Feb 15;188(4):1953-60. doi: 10.4049/jimmunol.1102543. Epub 2012 Jan 16. J Immunol. 2012. PMID: 22250086 Free PMC article.
-
T cell-intrinsic ASC critically promotes T(H)17-mediated experimental autoimmune encephalomyelitis.Nat Immunol. 2016 May;17(5):583-92. doi: 10.1038/ni.3389. Epub 2016 Mar 21. Nat Immunol. 2016. PMID: 26998763 Free PMC article.
-
Caspase-1/ASC inflammasome-mediated activation of IL-1β-ROS-NF-κB pathway for control of Trypanosoma cruzi replication and survival is dispensable in NLRP3-/- macrophages.PLoS One. 2014 Nov 5;9(11):e111539. doi: 10.1371/journal.pone.0111539. eCollection 2014. PLoS One. 2014. PMID: 25372293 Free PMC article.
-
Research progress on the NLRP3 inflammasome and its role in the central nervous system.Neurosci Bull. 2013 Dec;29(6):779-87. doi: 10.1007/s12264-013-1328-9. Epub 2013 Mar 20. Neurosci Bull. 2013. PMID: 23512739 Free PMC article. Review.
-
Negative regulation of NLRP3 inflammasome signaling.Protein Cell. 2013 Apr;4(4):251-8. doi: 10.1007/s13238-013-2128-8. Epub 2013 Mar 21. Protein Cell. 2013. PMID: 23519777 Free PMC article. Review.
Cited by
-
Interferon regulatory factor 4 plays a pivotal role in the development of aGVHD-associated colitis.Oncoimmunology. 2023 Dec 27;13(1):2296712. doi: 10.1080/2162402X.2023.2296712. eCollection 2024. Oncoimmunology. 2023. PMID: 38170159 Free PMC article.
-
Caspase-11 signaling enhances graft-versus-host disease.Nat Commun. 2019 Sep 6;10(1):4044. doi: 10.1038/s41467-019-11895-2. Nat Commun. 2019. PMID: 31492850 Free PMC article.
-
Targeting the NLRP3 inflammasome in inflammatory diseases.Nat Rev Drug Discov. 2018 Aug;17(8):588-606. doi: 10.1038/nrd.2018.97. Epub 2018 Jul 20. Nat Rev Drug Discov. 2018. PMID: 30026524 Review.
-
Cell type-specific roles of NLRP3, inflammasome-dependent and -independent, in host defense, sterile necroinflammation, tissue repair, and fibrosis.Front Immunol. 2023 Jul 25;14:1214289. doi: 10.3389/fimmu.2023.1214289. eCollection 2023. Front Immunol. 2023. PMID: 37564649 Free PMC article. Review.
-
Inflammatory Cytokine Networks in Gastrointestinal Tract Graft vs. Host Disease.Front Immunol. 2019 Feb 22;10:163. doi: 10.3389/fimmu.2019.00163. eCollection 2019. Front Immunol. 2019. PMID: 30853956 Free PMC article. Review.
References
-
- Akira S., Takeda K. 2004. Toll-like receptor signalling. Nat. Rev. Immunol. 4:499–511 http://www.nature.com/nri/journal/v4/n7/full/nri1391.html - PubMed
-
- Beelen D.W., Elmaagacli A., Müller K.D., Hirche H., Schaefer U.W. 1999. Influence of intestinal bacterial decontamination using metronidazole and ciprofloxacin or ciprofloxacin alone on the development of acute graft-versus-host disease after marrow transplantation in patients with hematologic malignancies: final results and long-term follow-up of an open-label prospective randomized trial. Blood. 93:3267–3275 - PubMed
-
- Bryson J.S., Zhang L., Goes S.W., Jennings C.D., Caywood B.E., Carlson S.L., Kaplan A.M. 2004. CD4+ T cells mediate murine syngeneic graft-versus-host disease-associated colitis. J. Immunol. 172:679–687 - PubMed
-
- Coccia M., Harrison O.J., Schiering C., Asquith M.J., Becher B., Powrie F., Maloy K.J. 2012. IL-1β mediates chronic intestinal inflammation by promoting the accumulation of IL-17A secreting innate lymphoid cells and CD4(+) Th17 cells. J. Exp. Med. 209:1595–1609 10.1084/jem.20111453 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
