

Auxin increases the hydrogen peroxide (H2O2) concentration in tomato (Solanum lycopersicum) root tips while inhibiting root growth
- PMID: 23965615
- PMCID: PMC3783245
- DOI: 10.1093/aob/mct181
Auxin increases the hydrogen peroxide (H2O2) concentration in tomato (Solanum lycopersicum) root tips while inhibiting root growth
Abstract
Background and aims: The hormone auxin and reactive oxygen species (ROS) regulate root elongation, but the interactions between the two pathways are not well understood. The aim of this study was to investigate how auxin interacts with ROS in regulating root elongation in tomato, Solanum lycopersicum.
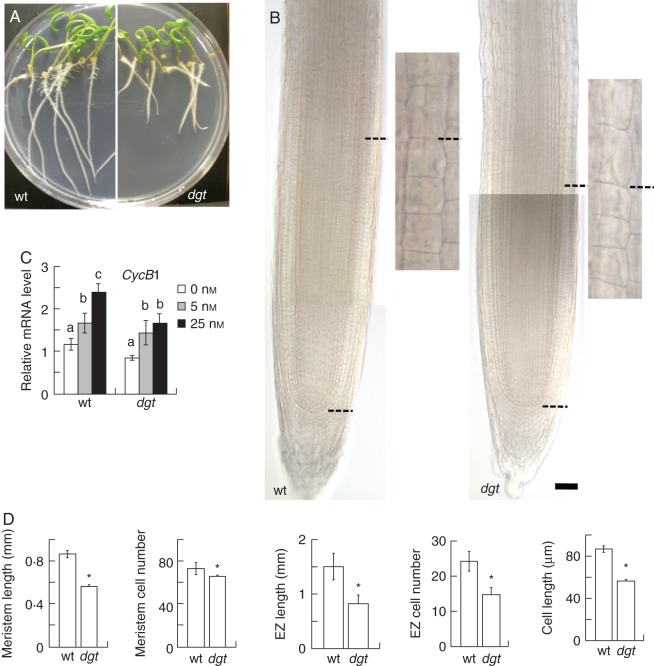
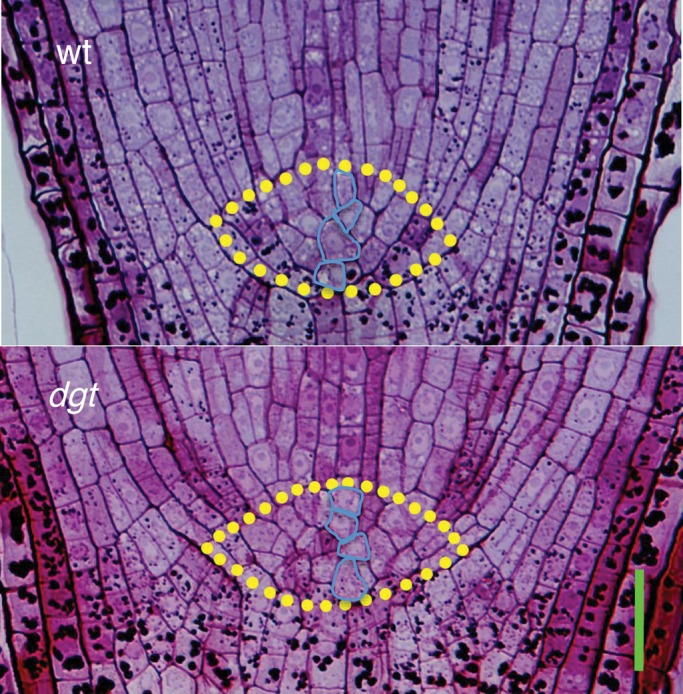
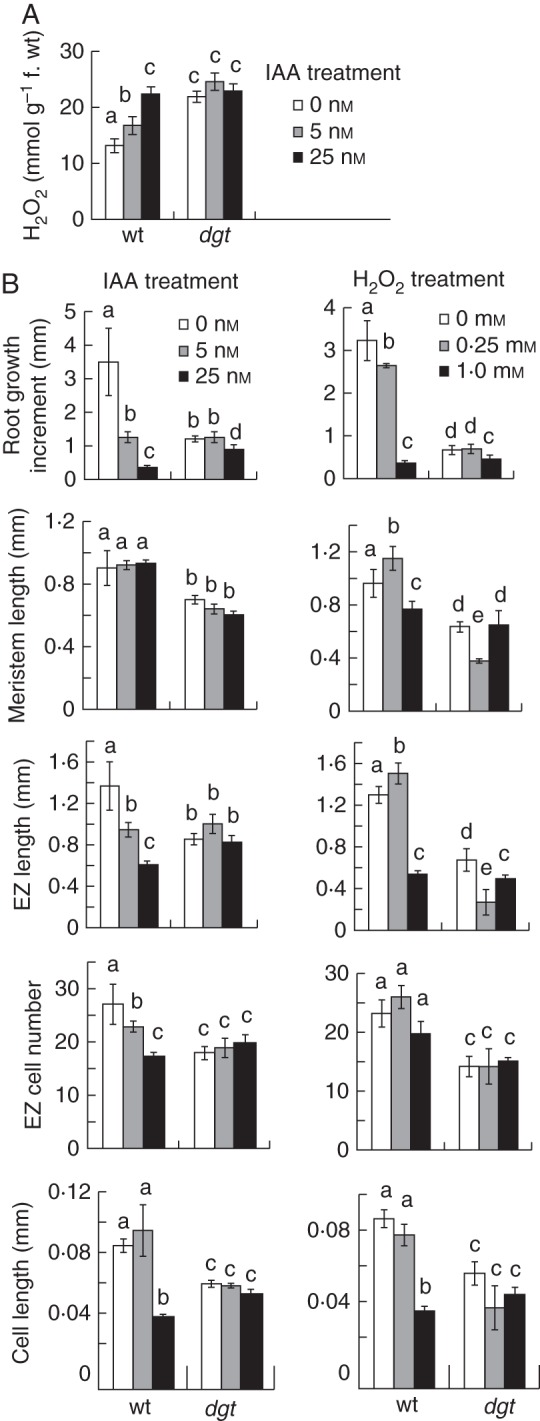
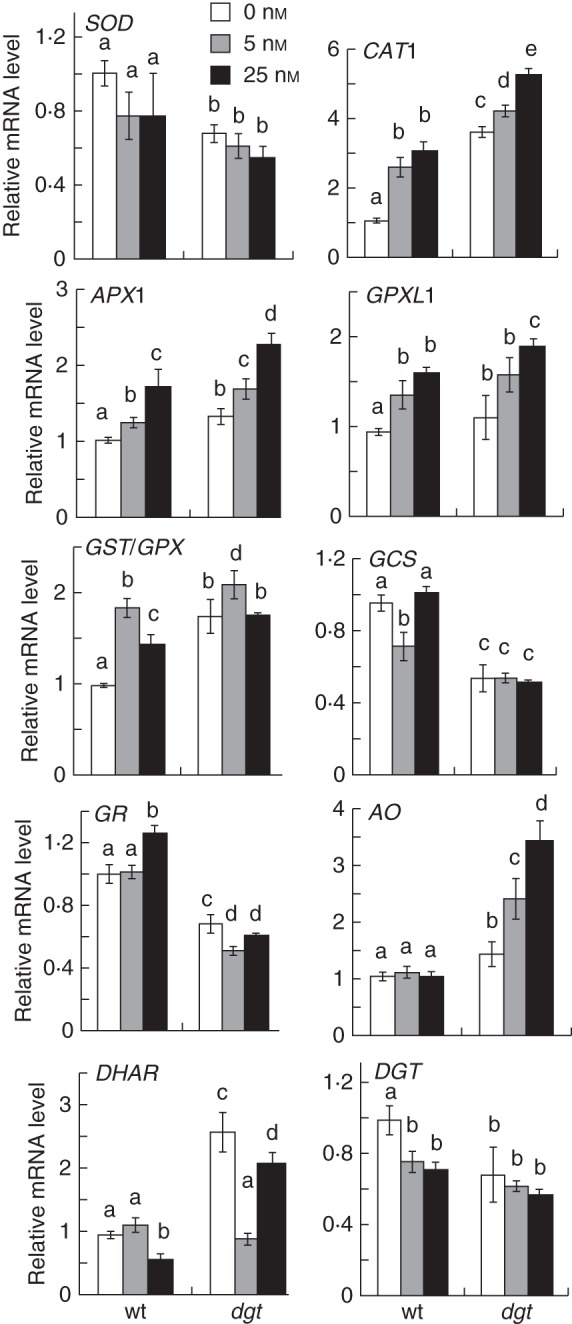
Methods: Wild-type and auxin-resistant mutant, diageotropica (dgt), of tomato (S. lycopersicum 'Ailsa Craig') were characterized in terms of root apical meristem and elongation zone histology, expression of the cell-cycle marker gene Sl-CycB1;1, accumulation of ROS, response to auxin and hydrogen peroxide (H2O2), and expression of ROS-related mRNAs.
Key results: The dgt mutant exhibited histological defects in the root apical meristem and elongation zone and displayed a constitutively increased level of hydrogen peroxide (H2O2) in the root tip, part of which was detected in the apoplast. Treatments of wild-type with auxin increased the H2O2 concentration in the root tip in a dose-dependent manner. Auxin and H2O2 elicited similar inhibition of cell elongation while bringing forth differential responses in terms of meristem length and number of cells in the elongation zone. Auxin treatments affected the expression of mRNAs of ROS-scavenging enzymes and less significantly mRNAs related to antioxidant level. The dgt mutation resulted in resistance to both auxin and H2O2 and affected profoundly the expression of mRNAs related to antioxidant level.
Conclusions: The results indicate that auxin regulates the level of H2O2 in the root tip, so increasing the auxin level triggers accumulation of H2O2 leading to inhibition of root cell elongation and root growth. The dgt mutation affects this pathway by reducing the auxin responsiveness of tissues and by disrupting the H2O2 homeostasis in the root tip.
Keywords: Auxin; ROS; Solanum lycopersicum; dgt; diageotropica; hydrogen peroxide; root elongation; tomato.
Figures


Similar articles
-
The diageotropica gene differentially affects auxin and cytokinin responses throughout development in tomato.Plant Physiol. 1998 May;117(1):63-72. doi: 10.1104/pp.117.1.63. Plant Physiol. 1998. PMID: 9576775 Free PMC article.
-
Involvement of reactive oxygen species and auxin in serotonin-induced inhibition of primary root elongation.J Plant Physiol. 2018 Oct;229:89-99. doi: 10.1016/j.jplph.2018.07.004. Epub 2018 Jul 18. J Plant Physiol. 2018. PMID: 30055520
-
Tomato fruit development in the auxin-resistant dgt mutant is induced by pollination but not by auxin treatment.J Plant Physiol. 2012 Aug 15;169(12):1165-72. doi: 10.1016/j.jplph.2012.04.005. Epub 2012 May 18. J Plant Physiol. 2012. PMID: 22608080
-
Just passing through: The auxin gradient of the root meristem.Curr Top Dev Biol. 2020;137:433-454. doi: 10.1016/bs.ctdb.2019.12.001. Epub 2020 Jan 7. Curr Top Dev Biol. 2020. PMID: 32143752 Review.
-
The root as a drill: an ethylene-auxin interaction facilitates root penetration in soil.Plant Signal Behav. 2012 Feb;7(2):151-6. doi: 10.4161/psb.18936. Epub 2012 Feb 1. Plant Signal Behav. 2012. PMID: 22415043 Free PMC article. Review.
Cited by
-
Hydrogen peroxide sulfenylates and inhibits the photorespiratory enzyme PGLP1 to modulate plant thermotolerance.Plant Commun. 2024 Jun 10;5(6):100852. doi: 10.1016/j.xplc.2024.100852. Epub 2024 Feb 25. Plant Commun. 2024. PMID: 38409783 Free PMC article.
-
The Role of Hydrogen Sulfide in Plant Roots during Development and in Response to Abiotic Stress.Int J Mol Sci. 2022 Jan 18;23(3):1024. doi: 10.3390/ijms23031024. Int J Mol Sci. 2022. PMID: 35162947 Free PMC article. Review.
-
Uncovering the Role of Hormones in Enhancing Antioxidant Defense Systems in Stressed Tomato (Solanum lycopersicum) Plants.Plants (Basel). 2023 Oct 23;12(20):3648. doi: 10.3390/plants12203648. Plants (Basel). 2023. PMID: 37896111 Free PMC article. Review.
-
NADPH Oxidase RbohD and Ethylene Signaling are Involved in Modulating Seedling Growth and Survival Under Submergence Stress.Plants (Basel). 2020 Apr 8;9(4):471. doi: 10.3390/plants9040471. Plants (Basel). 2020. PMID: 32276372 Free PMC article.
-
Ethanol Enhances High-Salinity Stress Tolerance by Detoxifying Reactive Oxygen Species in Arabidopsis thaliana and Rice.Front Plant Sci. 2017 Jul 3;8:1001. doi: 10.3389/fpls.2017.01001. eCollection 2017. Front Plant Sci. 2017. PMID: 28717360 Free PMC article.
References
-
- Barlow P. Stem cells and founder zones in plants, particularly their roots. In: Potten CS, editor. Stem cells. London: Academic Press; 1997. pp. 29–57.
-
- Baum SF, Rost TL. Root apical organization in Arabidopsis thaliana: 1. Root cap and protoderm. Protoplasma. 1996;192:178–188.
-
- Bielski BHJ, Shiue GG, Bajuk S. Reduction of nitro blue tetrazolium by CO2•- and O2•– radicals. Journal of Physiological Chemistry. 1980;84:830–833.
-
- Bienert GP, Møller AL, Kristiansen KA, et al. Specific aquaporins facilitate the diffusion of hydrogen peroxide across membranes. Journal of Biological Chemistry. 2007;282:1183–1192. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
