

The impact of age, biogenesis, and genomic clustering on Drosophila microRNA evolution
- PMID: 23882112
- PMCID: PMC3753935
- DOI: 10.1261/rna.039248.113
The impact of age, biogenesis, and genomic clustering on Drosophila microRNA evolution
Abstract
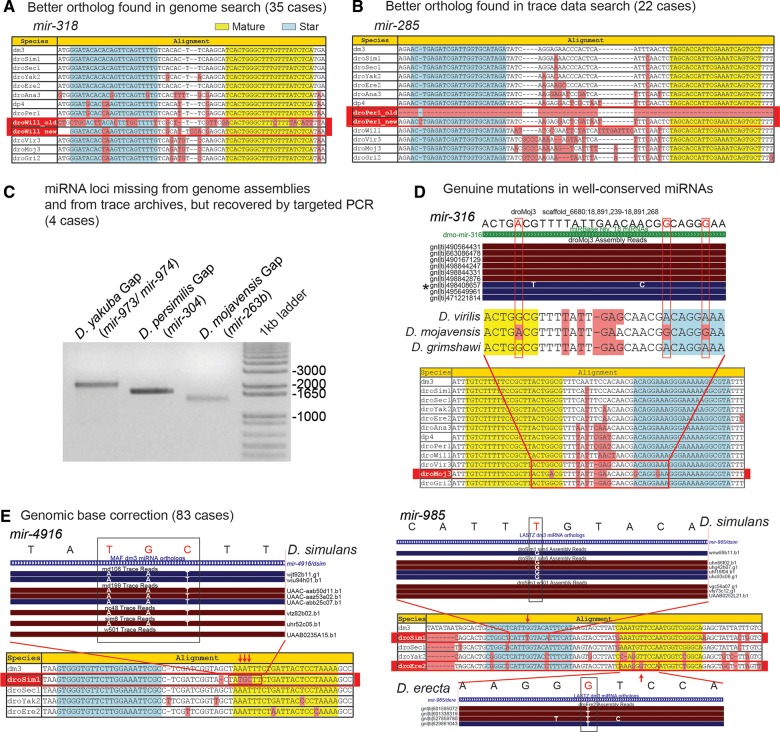
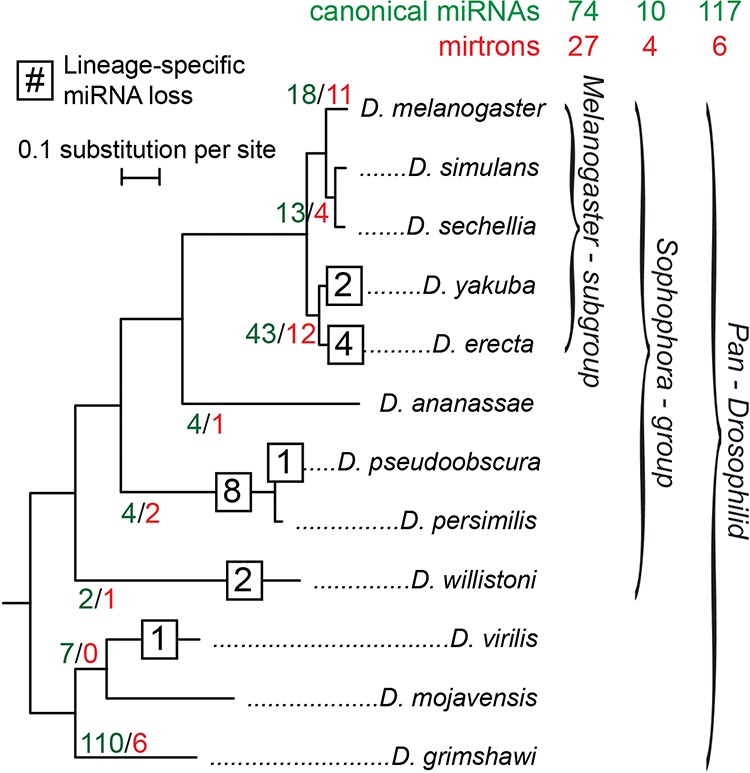
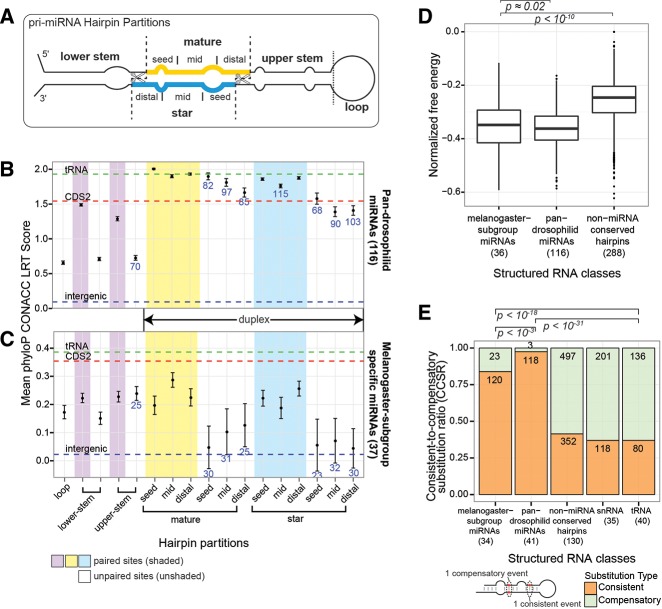
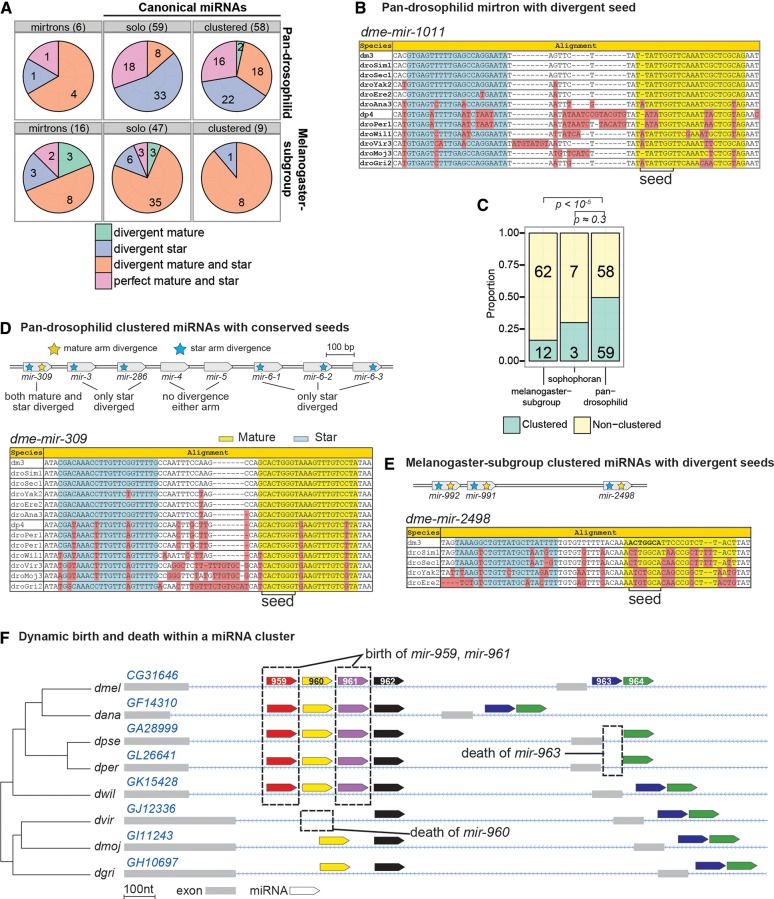
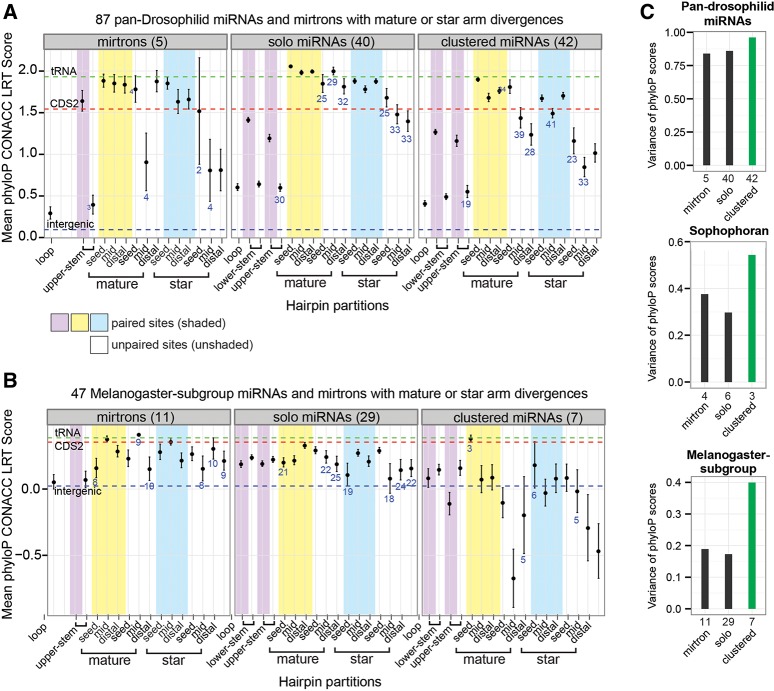
The molecular evolutionary signatures of miRNAs inform our understanding of their emergence, biogenesis, and function. The known signatures of miRNA evolution have derived mostly from the analysis of deeply conserved, canonical loci. In this study, we examine the impact of age, biogenesis pathway, and genomic arrangement on the evolutionary properties of Drosophila miRNAs. Crucial to the accuracy of our results was our curation of high-quality miRNA alignments, which included nearly 150 corrections to ortholog calls and nucleotide sequences of the global 12-way Drosophilid alignments currently available. Using these data, we studied primary sequence conservation, normalized free-energy values, and types of structure-preserving substitutions. We expand upon common miRNA evolutionary patterns that reflect fundamental features of miRNAs that are under functional selection. We observe that melanogaster-subgroup-specific miRNAs, although recently emerged and rapidly evolving, nonetheless exhibit evolutionary signatures that are similar to well-conserved miRNAs and distinct from other structured noncoding RNAs and bulk conserved non-miRNA hairpins. This provides evidence that even young miRNAs may be selected for regulatory activities. More strikingly, we observe that mirtrons and clustered miRNAs both exhibit distinct evolutionary properties relative to solo, well-conserved miRNAs, even after controlling for sequence depth. These studies highlight the previously unappreciated impact of biogenesis strategy and genomic location on the evolutionary dynamics of miRNAs, and affirm that miRNAs do not evolve as a unitary class.
Keywords: Drosophila; microRNA clusters; microRNA evolution; mirtrons; structure evolution.
Figures
Similar articles
-
Adaptive evolution of testis-specific, recently evolved, clustered miRNAs in Drosophila.RNA. 2014 Aug;20(8):1195-209. doi: 10.1261/rna.044644.114. Epub 2014 Jun 18. RNA. 2014. PMID: 24942624 Free PMC article.
-
Computational identification of Drosophila microRNA genes.Genome Biol. 2003;4(7):R42. doi: 10.1186/gb-2003-4-7-r42. Epub 2003 Jun 30. Genome Biol. 2003. PMID: 12844358 Free PMC article.
-
Diverse modes of evolutionary emergence and flux of conserved microRNA clusters.RNA. 2014 Dec;20(12):1850-63. doi: 10.1261/rna.046805.114. Epub 2014 Oct 20. RNA. 2014. PMID: 25332374 Free PMC article.
-
Regulatory RNAs in the light of Drosophila genomics.Brief Funct Genomics. 2012 Sep;11(5):356-65. doi: 10.1093/bfgp/els033. Epub 2012 Sep 5. Brief Funct Genomics. 2012. PMID: 22956639 Free PMC article. Review.
-
Convergent and divergent evolution of microRNA-mediated regulation in metazoans.Biol Rev Camb Philos Soc. 2024 Apr;99(2):525-545. doi: 10.1111/brv.13033. Epub 2023 Nov 21. Biol Rev Camb Philos Soc. 2024. PMID: 37987240 Review.
Cited by
-
The Drosophila Dicer-1 Partner Loquacious Enhances miRNA Processing from Hairpins with Unstable Structures at the Dicing Site.Cell Rep. 2016 May 24;15(8):1795-808. doi: 10.1016/j.celrep.2016.04.059. Epub 2016 May 12. Cell Rep. 2016. PMID: 27184838 Free PMC article.
-
Predicting microRNA targeting efficacy in Drosophila.Genome Biol. 2018 Oct 4;19(1):152. doi: 10.1186/s13059-018-1504-3. Genome Biol. 2018. PMID: 30286781 Free PMC article.
-
Evolutionary Comparison of Two Combinatorial Regulators of SBP-Box Genes, MiR156 and MiR529, in Plants.PLoS One. 2015 Apr 24;10(4):e0124621. doi: 10.1371/journal.pone.0124621. eCollection 2015. PLoS One. 2015. PMID: 25909360 Free PMC article.
-
Parameters of clustered suboptimal miRNA biogenesis.Proc Natl Acad Sci U S A. 2023 Oct 10;120(41):e2306727120. doi: 10.1073/pnas.2306727120. Epub 2023 Oct 3. Proc Natl Acad Sci U S A. 2023. PMID: 37788316 Free PMC article.
-
Identification and consequences of miRNA-target interactions--beyond repression of gene expression.Nat Rev Genet. 2014 Sep;15(9):599-612. doi: 10.1038/nrg3765. Epub 2014 Jul 15. Nat Rev Genet. 2014. PMID: 25022902 Review.
References
-
- Aravin A, Lagos-Quintana M, Yalcin A, Zavolan M, Marks D, Snyder B, Gaasterland T, Meyer J, Tuschl T 2003. The small RNA profile during Drosophila melanogaster development. Dev Cell 5: 337–350 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases