

Anle138b: a novel oligomer modulator for disease-modifying therapy of neurodegenerative diseases such as prion and Parkinson's disease
- PMID: 23604588
- PMCID: PMC3661926
- DOI: 10.1007/s00401-013-1114-9
Anle138b: a novel oligomer modulator for disease-modifying therapy of neurodegenerative diseases such as prion and Parkinson's disease
Abstract
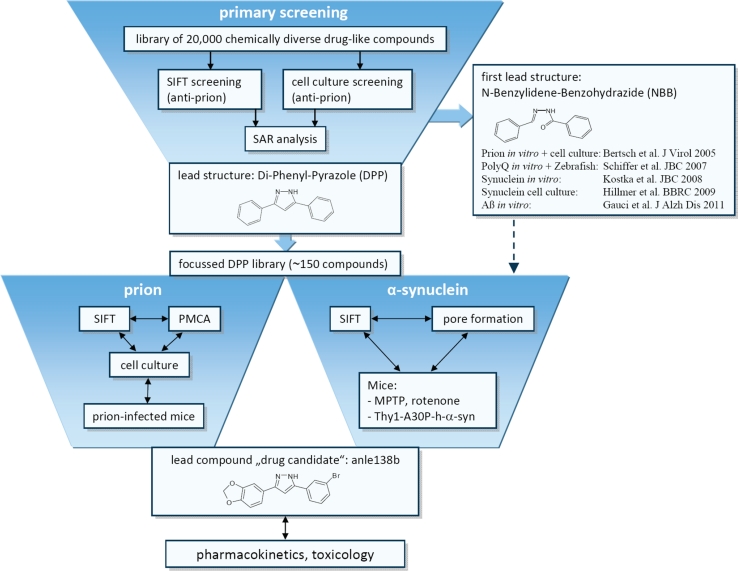
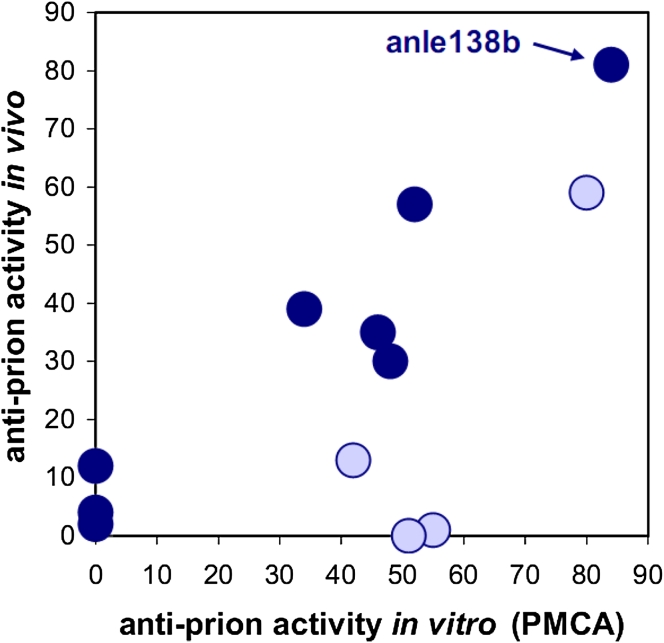
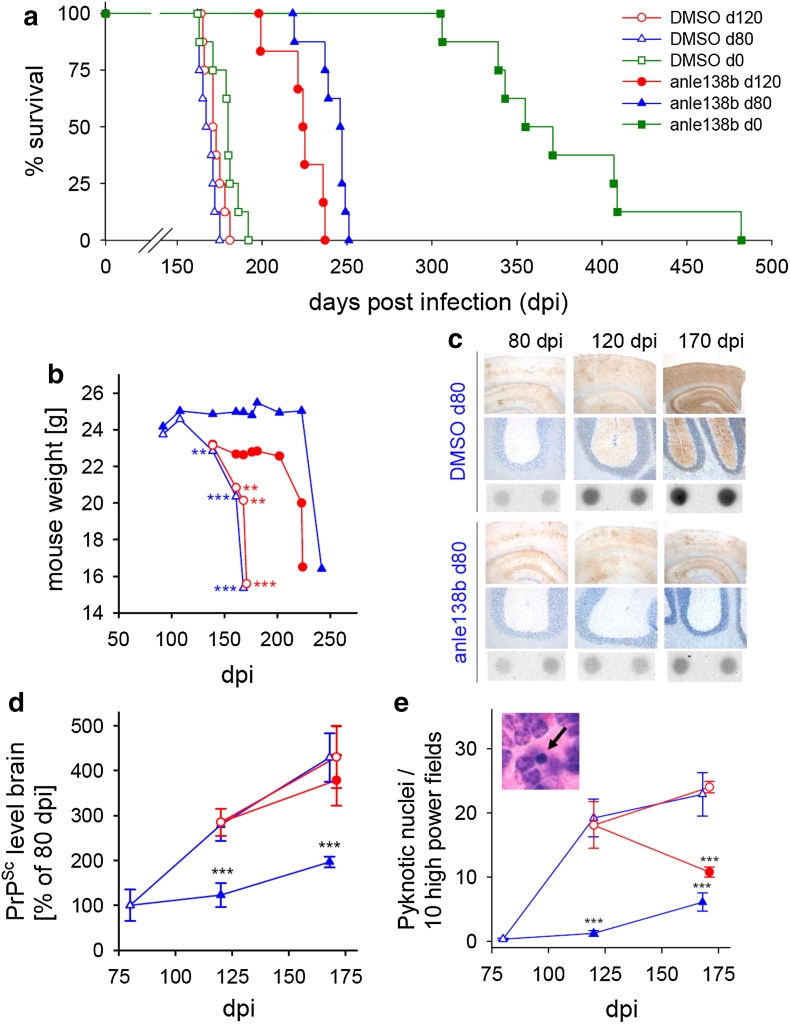
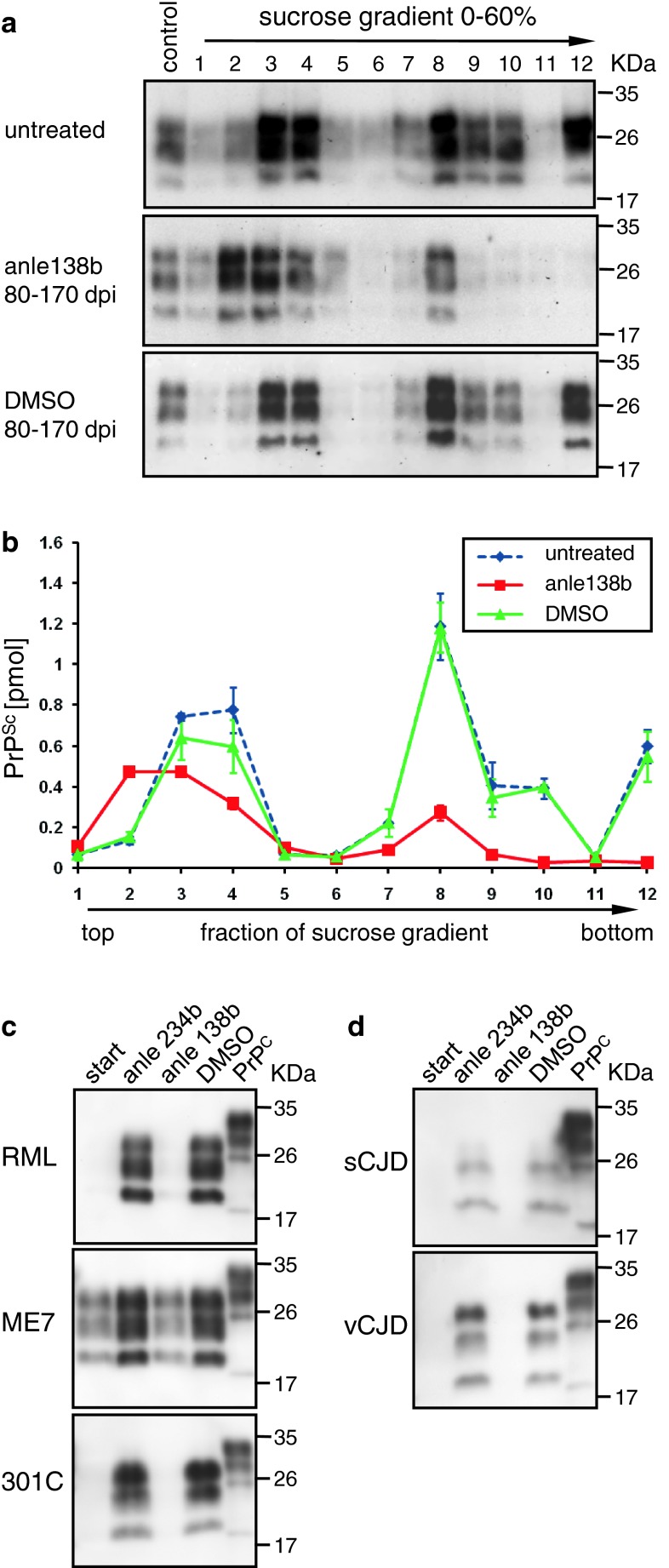
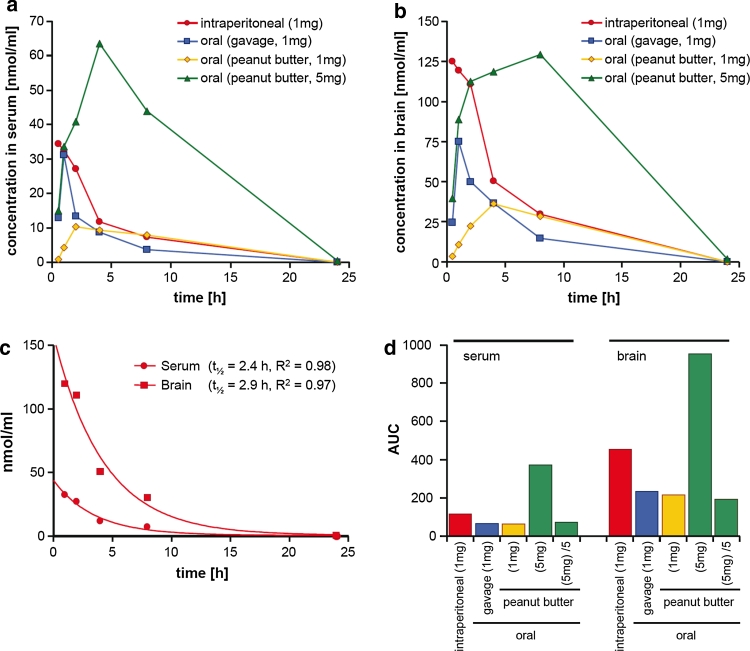
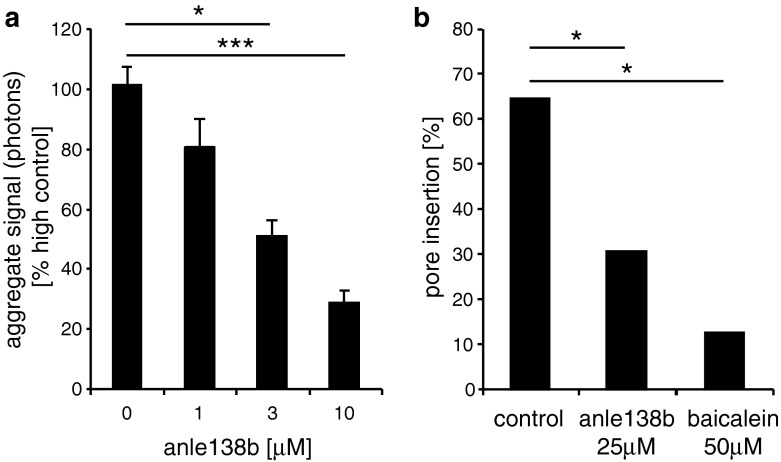
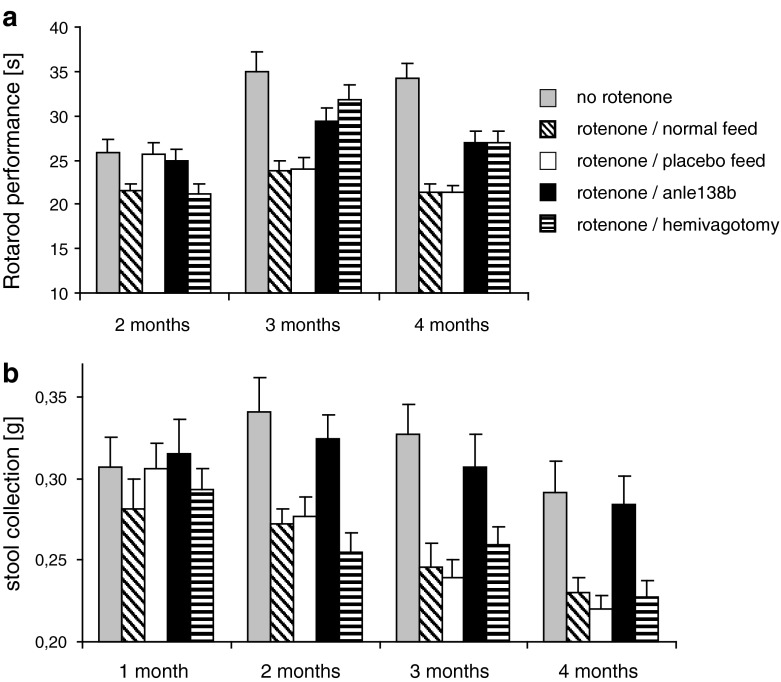
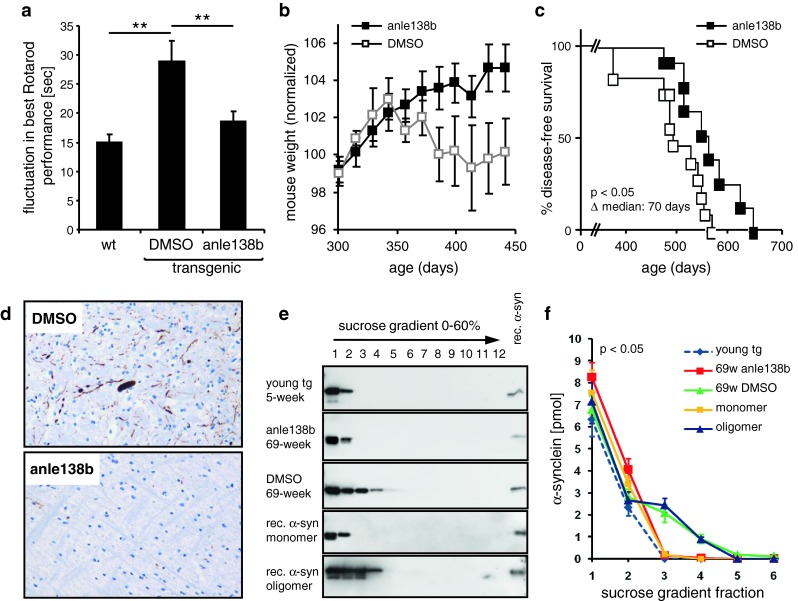
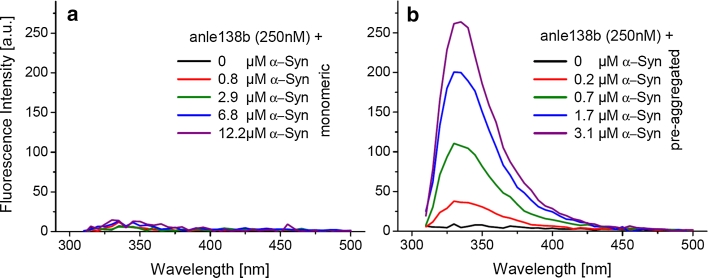
In neurodegenerative diseases such as Alzheimer's disease (AD), Parkinson's disease (PD) and prion diseases, deposits of aggregated disease-specific proteins are found. Oligomeric aggregates are presumed to be the key neurotoxic agent. Here we describe the novel oligomer modulator anle138b [3-(1,3-benzodioxol-5-yl)-5-(3-bromophenyl)-1H-pyrazole], an aggregation inhibitor we developed based on a systematic high-throughput screening campaign combined with medicinal chemistry optimization. In vitro, anle138b blocked the formation of pathological aggregates of prion protein (PrP(Sc)) and of α-synuclein (α-syn), which is deposited in PD and other synucleinopathies such as dementia with Lewy bodies (DLB) and multiple system atrophy (MSA). Notably, anle138b strongly inhibited all prion strains tested including BSE-derived and human prions. Anle138b showed structure-dependent binding to pathological aggregates and strongly inhibited formation of pathological oligomers in vitro and in vivo both for prion protein and α-synuclein. Both in mouse models of prion disease and in three different PD mouse models, anle138b strongly inhibited oligomer accumulation, neuronal degeneration, and disease progression in vivo. Anle138b had no detectable toxicity at therapeutic doses and an excellent oral bioavailability and blood-brain-barrier penetration. Our findings indicate that oligomer modulators provide a new approach for disease-modifying therapy in these diseases, for which only symptomatic treatment is available so far. Moreover, our findings suggest that pathological oligomers in neurodegenerative diseases share structural features, although the main protein component is disease-specific, indicating that compounds such as anle138b that modulate oligomer formation by targeting structure-dependent epitopes can have a broad spectrum of activity in the treatment of different protein aggregation diseases.
Figures
Similar articles
-
Safety, tolerability and pharmacokinetics of the oligomer modulator anle138b with exposure levels sufficient for therapeutic efficacy in a murine Parkinson model: A randomised, double-blind, placebo-controlled phase 1a trial.EBioMedicine. 2022 Jun;80:104021. doi: 10.1016/j.ebiom.2022.104021. Epub 2022 Apr 29. EBioMedicine. 2022. PMID: 35500536 Free PMC article. Clinical Trial.
-
Anle138b and related compounds are aggregation specific fluorescence markers and reveal high affinity binding to α-synuclein aggregates.Biochim Biophys Acta. 2015 Sep;1850(9):1884-90. doi: 10.1016/j.bbagen.2015.05.021. Epub 2015 May 29. Biochim Biophys Acta. 2015. PMID: 26028294
-
The prion hypothesis of Parkinson's disease.Curr Neurol Neurosci Rep. 2015 May;15(5):28. doi: 10.1007/s11910-015-0549-x. Curr Neurol Neurosci Rep. 2015. PMID: 25868519
-
How strong is the evidence that Parkinson's disease is a prion disorder?Curr Opin Neurol. 2016 Aug;29(4):459-66. doi: 10.1097/WCO.0000000000000349. Curr Opin Neurol. 2016. PMID: 27257944 Free PMC article. Review.
-
Structural and Functional Insights into α-Synuclein Fibril Polymorphism.Biomolecules. 2021 Sep 28;11(10):1419. doi: 10.3390/biom11101419. Biomolecules. 2021. PMID: 34680054 Free PMC article. Review.
Cited by
-
Safety, tolerability and pharmacokinetics of the oligomer modulator anle138b with exposure levels sufficient for therapeutic efficacy in a murine Parkinson model: A randomised, double-blind, placebo-controlled phase 1a trial.EBioMedicine. 2022 Jun;80:104021. doi: 10.1016/j.ebiom.2022.104021. Epub 2022 Apr 29. EBioMedicine. 2022. PMID: 35500536 Free PMC article. Clinical Trial.
-
Therapeutic strategies for Parkinson disease: beyond dopaminergic drugs.Nat Rev Drug Discov. 2018 Nov;17(11):804-822. doi: 10.1038/nrd.2018.136. Epub 2018 Sep 28. Nat Rev Drug Discov. 2018. PMID: 30262889 Review.
-
Small molecule inhibits α-synuclein aggregation, disrupts amyloid fibrils, and prevents degeneration of dopaminergic neurons.Proc Natl Acad Sci U S A. 2018 Oct 9;115(41):10481-10486. doi: 10.1073/pnas.1804198115. Epub 2018 Sep 24. Proc Natl Acad Sci U S A. 2018. PMID: 30249646 Free PMC article.
-
Alpha-Synuclein: From Early Synaptic Dysfunction to Neurodegeneration.Front Neurol. 2018 May 4;9:295. doi: 10.3389/fneur.2018.00295. eCollection 2018. Front Neurol. 2018. PMID: 29780350 Free PMC article. Review.
-
α-Helical peptidic scaffolds to target α-synuclein toxic species with nanomolar affinity.Nat Commun. 2021 Jun 18;12(1):3752. doi: 10.1038/s41467-021-24039-2. Nat Commun. 2021. PMID: 34145261 Free PMC article.
References
-
- Bertsch U, Winklhofer KF, Hirschberger T, Bieschke J, Weber P, Hartl FU, Tavan P, Tatzelt J, Kretzschmar HA, Giese A. Systematic identification of antiprion drugs by high-throughput screening based on scanning for intensely fluorescent targets. J Virol. 2005;79:7785–7791. doi: 10.1128/JVI.79.12.7785-7791.2005. - DOI - PMC - PubMed
-
- Bieschke J, Herbst M, Wiglenda T, Friedrich RP, Boeddrich A, Schiele F, Kleckers D, Lopez del Amo JM, Gruning BA, Wang Q, Schmidt MR, Lurz R, Anwyl R, Schnoegl S, Fandrich M, Frank RF, Reif B, Gunther S, Walsh DM, Wanker EE. Small-molecule conversion of toxic oligomers to nontoxic beta-sheet-rich amyloid fibrils. Nat Chem Biol. 2011;8:93–101. doi: 10.1038/nchembio.719. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
