

Quantification of growth-defense trade-offs in a common currency: nitrogen required for phenolamide biosynthesis is not derived from ribulose-1,5-bisphosphate carboxylase/oxygenase turnover
- PMID: 23590461
- PMCID: PMC4996319
- DOI: 10.1111/tpj.12210
Quantification of growth-defense trade-offs in a common currency: nitrogen required for phenolamide biosynthesis is not derived from ribulose-1,5-bisphosphate carboxylase/oxygenase turnover
Abstract
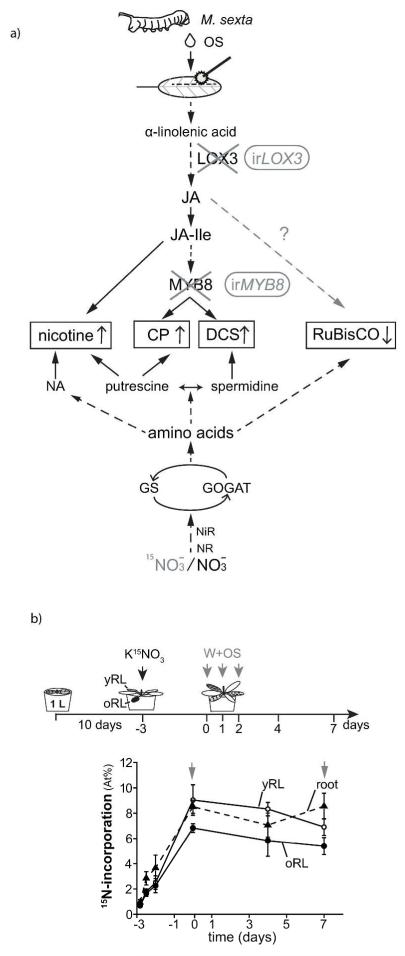
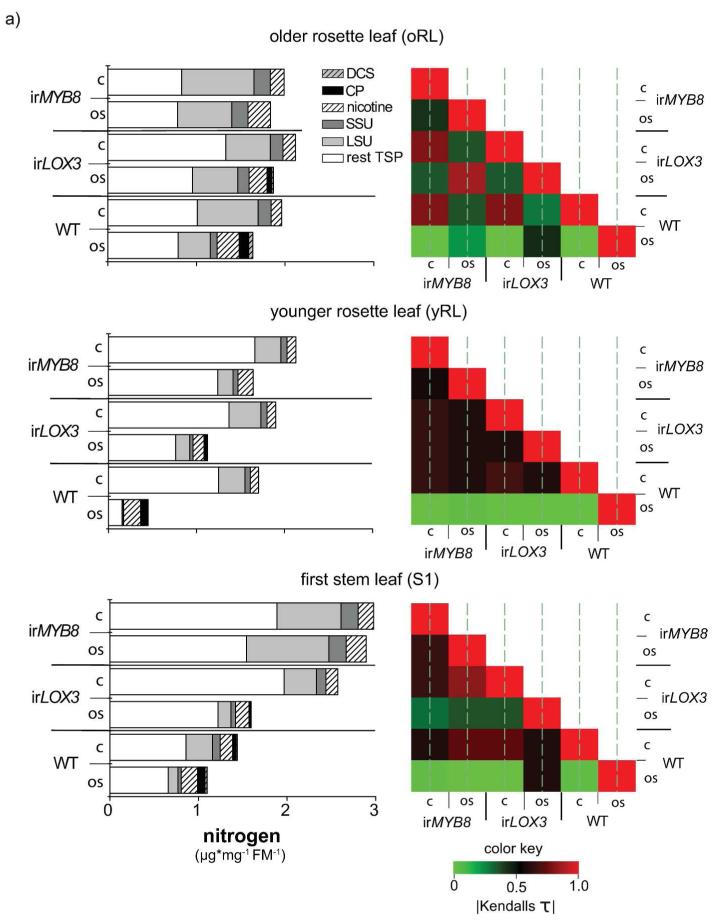
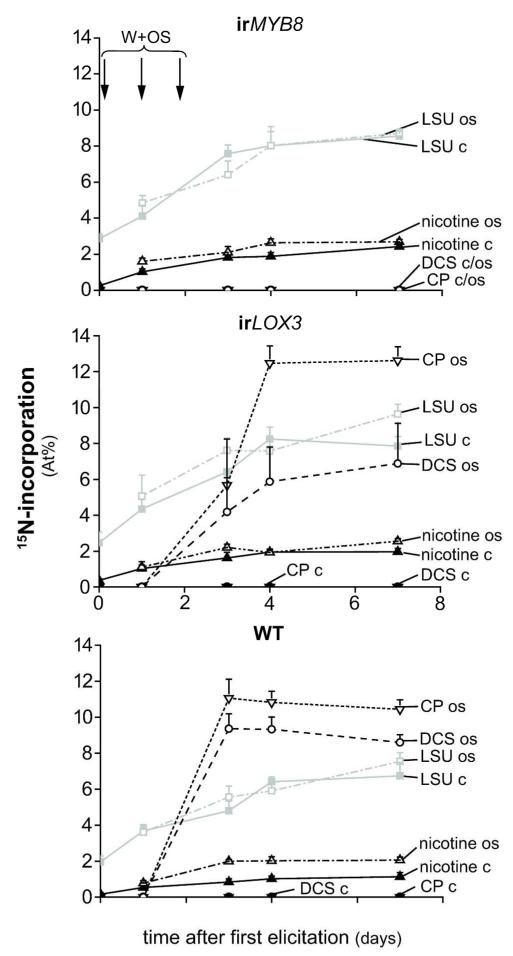
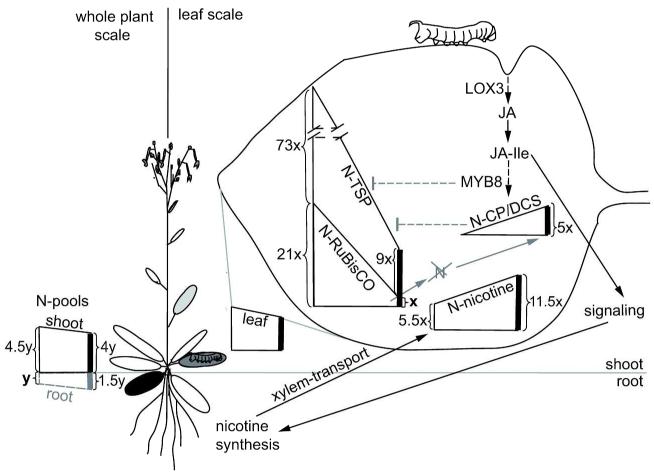
Induced defenses are thought to be economical: growth and fitness-limiting resources are only invested into defenses when needed. To date, this putative growth-defense trade-off has not been quantified in a common currency at the level of individual compounds. Here, a quantification method for ¹⁵N-labeled proteins enabled a direct comparison of nitrogen (N) allocation to proteins, specifically, ribulose-1,5-bisposphate carboxylase/oxygenase (RuBisCO), as proxy for growth, with that to small N-containing defense metabolites (nicotine and phenolamides), as proxies for defense after herbivory. After repeated simulated herbivory, total N decreased in the shoots of wild-type (WT) Nicotiana attenuata plants, but not in two transgenic lines impaired in jasmonate defense signaling (irLOX3) and phenolamide biosynthesis (irMYB8). N was reallocated among different compounds within elicited rosette leaves: in the WT, a strong decrease in total soluble protein (TSP) and RuBisCO was accompanied by an increase in defense metabolites, irLOX3 showed a similar, albeit attenuated, pattern, whereas irMYB8 rosette leaves were the least responsive to elicitation, with overall higher levels of RuBisCO. Induced defenses were higher in the older compared with the younger rosette leaves, supporting the hypothesis that tissue developmental stage influences defense investments. We propose that MYB8, probably by regulating the production of phenolamides, indirectly mediates protein pool sizes after herbivory. Although the decrease in absolute N invested in TSP and RuBisCO elicited by simulated herbivory was much larger than the N-requirements of nicotine and phenolamide biosynthesis, ¹⁵N flux studies revealed that N for phenolamide synthesis originates from recently assimilated N, rather than from RuBisCO turnover.
Keywords: Manduca sexta; Nicotiana attenuata; R2R3-MYB transcription factor; caffeoyl-putrescine; dicaffeoyl-spermidine; nicotine; ribulose-1,5-bisphosphate carboxylase/oxygenase; total soluble protein.
© 2013 The Authors The Plant Journal © 2013 John Wiley & Sons Ltd.
Figures



Similar articles
-
Silencing ribulose-1,5-bisphosphate carboxylase/oxygenase expression does not disrupt nitrogen allocation to defense after simulated herbivory in Nicotiana attenuata.Plant Signal Behav. 2013;8(12):e27570. doi: 10.4161/psb.27570. Epub 2013 Dec 31. Plant Signal Behav. 2013. PMID: 24390158 Free PMC article.
-
Ethylene is a local modulator of jasmonate-dependent phenolamide accumulation during Manduca sexta herbivory in Nicotiana attenuata.Plant Cell Environ. 2021 Mar;44(3):964-981. doi: 10.1111/pce.13955. Epub 2020 Dec 15. Plant Cell Environ. 2021. PMID: 33215737
-
MAPK-dependent JA and SA signalling in Nicotiana attenuata affects plant growth and fitness during competition with conspecifics.BMC Plant Biol. 2012 Nov 13;12:213. doi: 10.1186/1471-2229-12-213. BMC Plant Biol. 2012. PMID: 23148462 Free PMC article.
-
The essential role of jasmonic acid in plant-herbivore interactions--using the wild tobacco Nicotiana attenuata as a model.J Genet Genomics. 2013 Dec 20;40(12):597-606. doi: 10.1016/j.jgg.2013.10.001. Epub 2013 Nov 9. J Genet Genomics. 2013. PMID: 24377866 Review.
-
Revealing insect herbivory-induced phenolamide metabolism: from single genes to metabolic network plasticity analysis.Plant J. 2014 Aug;79(4):679-92. doi: 10.1111/tpj.12503. Epub 2014 May 21. Plant J. 2014. PMID: 24617849 Free PMC article. Review.
Cited by
-
A Phytochrome B-Independent Pathway Restricts Growth at High Levels of Jasmonate Defense.Plant Physiol. 2020 Jun;183(2):733-749. doi: 10.1104/pp.19.01335. Epub 2020 Apr 3. Plant Physiol. 2020. PMID: 32245790 Free PMC article.
-
Metatranscriptomic Comparison of Endophytic and Pathogenic Fusarium-Arabidopsis Interactions Reveals Plant Transcriptional Plasticity.Mol Plant Microbe Interact. 2021 Sep;34(9):1071-1083. doi: 10.1094/MPMI-03-21-0063-R. Epub 2021 Oct 11. Mol Plant Microbe Interact. 2021. PMID: 33856230 Free PMC article.
-
Functional variation in a key defense gene structures herbivore communities and alters plant performance.PLoS One. 2018 Jun 6;13(6):e0197221. doi: 10.1371/journal.pone.0197221. eCollection 2018. PLoS One. 2018. PMID: 29874269 Free PMC article.
-
Plant-Growth-Promoting Rhizobacteria Modulate Carbohydrate Metabolism in Connection with Host Plant Defense Mechanism.Int J Mol Sci. 2024 Jan 25;25(3):1465. doi: 10.3390/ijms25031465. Int J Mol Sci. 2024. PMID: 38338742 Free PMC article. Review.
-
Changes in cytokinins are sufficient to alter developmental patterns of defense metabolites in Nicotiana attenuata.Plant J. 2017 Jan;89(1):15-30. doi: 10.1111/tpj.13316. Plant J. 2017. PMID: 27557345 Free PMC article.
References
-
- Allmann S, Halitschke R, Schuurink RC, Baldwin IT. Oxylipin channelling in Nicotiana attenuata: lipoxygenase 2 supplies substrates for green leaf volatile production. Plant Cell Environ. 2010;33:2028–2040. - PubMed
-
- Baldwin IT. Inducible nicotine production in native Nicotiana as an example of adaptive phenotypic plasticity. J. Chem. Ecol. 1999;25:3–30.
-
- Baldwin IT, Gorham D, Schmelz EA, Lewandowski CA, Lynds GY. Allocation of nitrogen to an inducible defense and seed production in Nicotiana attenuata. Oecologia. 1998;115:541–552. - PubMed
-
- Baldwin IT, Hamilton W. Jasmonate-induced responses of Nicotiana sylvestris results in fitness costs due to impaired competitive ability for nitrogen. J. Chem. Ecol. 2000;26:915–952.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
