

Embryonic mammary signature subsets are activated in Brca1-/- and basal-like breast cancers
- PMID: 23506684
- PMCID: PMC3672751
- DOI: 10.1186/bcr3403
Embryonic mammary signature subsets are activated in Brca1-/- and basal-like breast cancers
Abstract
Introduction: Cancer is often suggested to result from development gone awry. Links between normal embryonic development and cancer biology have been postulated, but no defined genetic basis has been established. We recently published the first transcriptomic analysis of embryonic mammary cell populations. Embryonic mammary epithelial cells are an immature progenitor cell population, lacking differentiation markers, which is reflected in their very distinct genetic profiles when compared with those of their postnatal descendents.
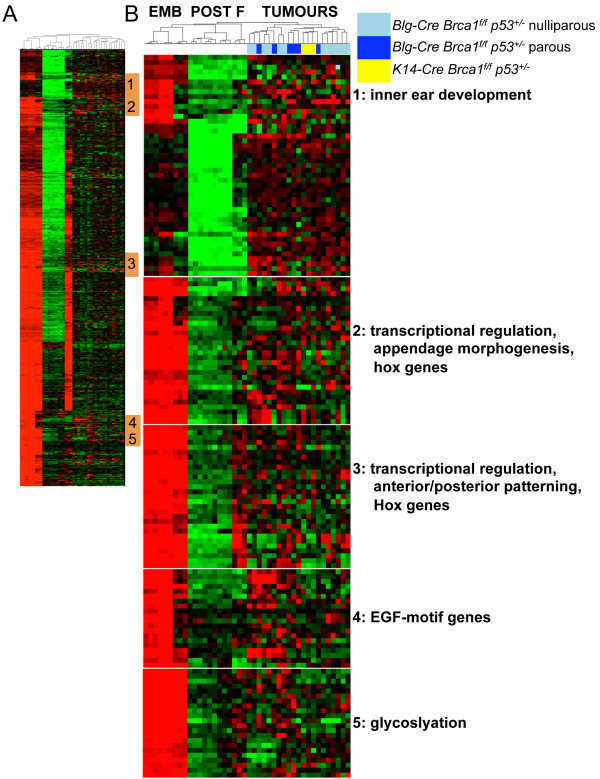
Methods: We defined an embryonic mammary epithelial signature that incorporates the most highly expressed genes from embryonic mammary epithelium when compared with the postnatal mammary epithelial cells. We looked for activation of the embryonic mammary epithelial signature in mouse mammary tumors that formed in mice in which Brca1 had been conditionally deleted from the mammary epithelium and in human breast cancers to determine whether any genetic links exist between embryonic mammary cells and breast cancers.
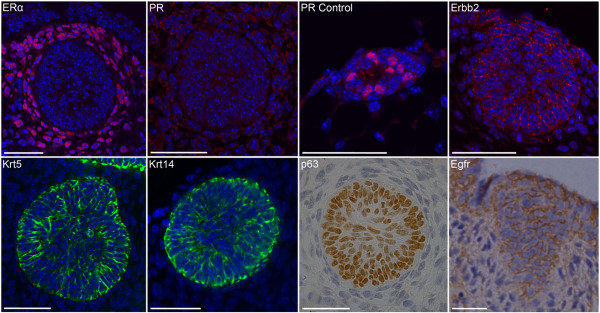
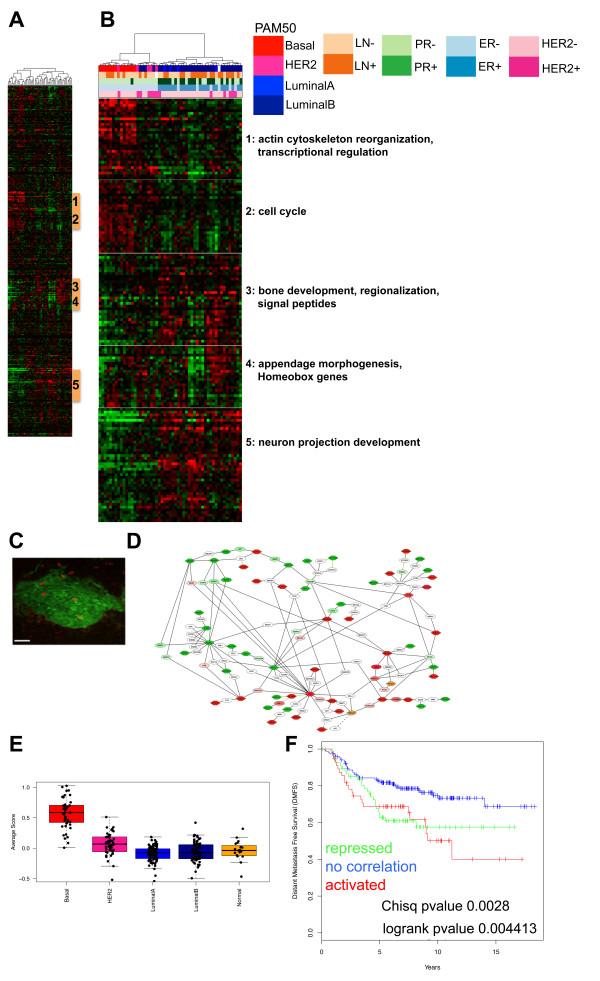
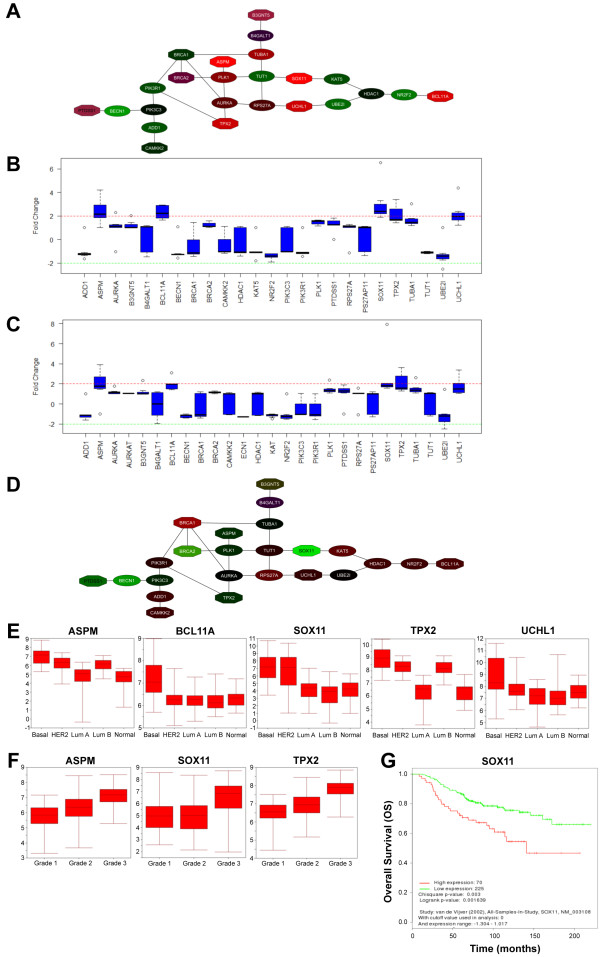
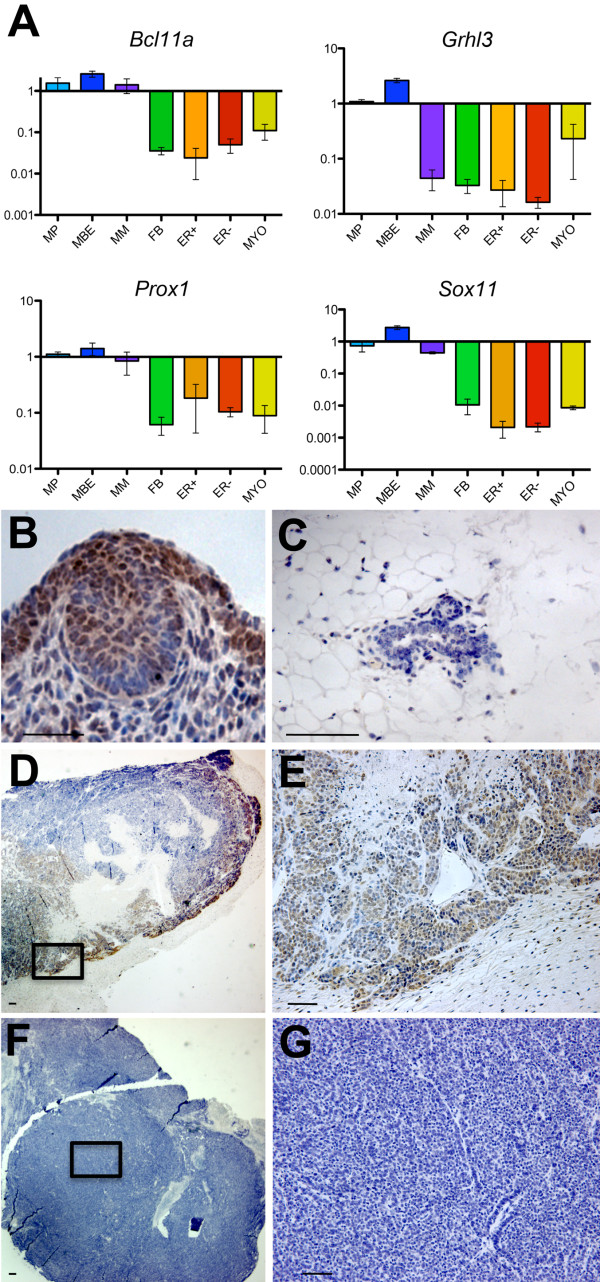
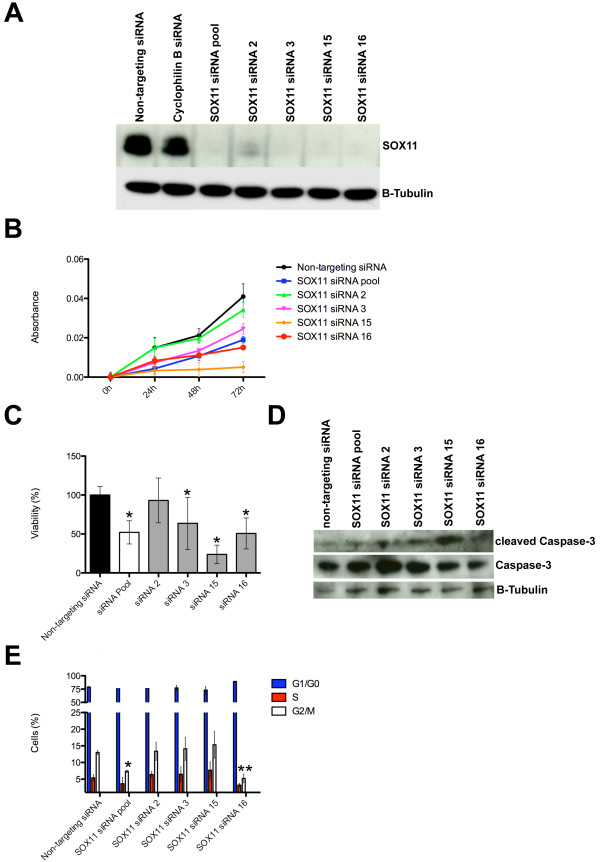
Results: Small subsets of the embryonic mammary epithelial signature were consistently activated in mouse Brca1-/- tumors and human basal-like breast cancers, which encoded predominantly transcriptional regulators, cell-cycle, and actin cytoskeleton components. Other embryonic gene subsets were found activated in non-basal-like tumor subtypes and repressed in basal-like tumors, including regulators of neuronal differentiation, transcription, and cell biosynthesis. Several embryonic genes showed significant upregulation in estrogen receptor (ER)-negative, progesterone receptor (PR)-negative, and/or grade 3 breast cancers. Among them, the transcription factor, SOX11, a progenitor cell and lineage regulator of nonmammary cell types, is found highly expressed in some Brca1-/- mammary tumors. By using RNA interference to silence SOX11 expression in breast cancer cells, we found evidence that SOX11 regulates breast cancer cell proliferation and cell survival.
Conclusions: Specific subsets of embryonic mammary genes, rather than the entire embryonic development transcriptomic program, are activated in tumorigenesis. Genes involved in embryonic mammary development are consistently upregulated in some breast cancers and warrant further investigation, potentially in drug-discovery research endeavors.
Figures
Similar articles
-
c-Kit is required for growth and survival of the cells of origin of Brca1-mutation-associated breast cancer.Oncogene. 2012 Feb 16;31(7):869-83. doi: 10.1038/onc.2011.289. Epub 2011 Jul 18. Oncogene. 2012. PMID: 21765473
-
Parity induces differentiation and reduces Wnt/Notch signaling ratio and proliferation potential of basal stem/progenitor cells isolated from mouse mammary epithelium.Breast Cancer Res. 2013 Apr 29;15(2):R36. doi: 10.1186/bcr3419. Breast Cancer Res. 2013. PMID: 23621987 Free PMC article.
-
Loss of BRCA1 leads to an increase in epidermal growth factor receptor expression in mammary epithelial cells, and epidermal growth factor receptor inhibition prevents estrogen receptor-negative cancers in BRCA1-mutant mice.Breast Cancer Res. 2011 Mar 11;13(2):R30. doi: 10.1186/bcr2850. Breast Cancer Res. 2011. PMID: 21396117 Free PMC article.
-
The cell of origin of BRCA1 mutation-associated breast cancer: a cautionary tale of gene expression profiling.J Mammary Gland Biol Neoplasia. 2011 Apr;16(1):51-5. doi: 10.1007/s10911-011-9202-8. Epub 2011 Feb 19. J Mammary Gland Biol Neoplasia. 2011. PMID: 21336547 Review.
-
A mouse mammary gland involution mRNA signature identifies biological pathways potentially associated with breast cancer metastasis.J Mammary Gland Biol Neoplasia. 2009 Jun;14(2):99-116. doi: 10.1007/s10911-009-9120-1. Epub 2009 Apr 30. J Mammary Gland Biol Neoplasia. 2009. PMID: 19408105 Review.
Cited by
-
Secondary Analysis of Human Bulk RNA-Seq Dataset Suggests Potential Mechanisms for Letrozole Resistance in Estrogen-Positive (ER+) Breast Cancer.Curr Issues Mol Biol. 2024 Jul 6;46(7):7114-7133. doi: 10.3390/cimb46070424. Curr Issues Mol Biol. 2024. PMID: 39057065 Free PMC article.
-
A human organoid system that self-organizes to recapitulate growth and differentiation of a benign mammary tumor.Proc Natl Acad Sci U S A. 2019 Jun 4;116(23):11444-11453. doi: 10.1073/pnas.1702372116. Epub 2019 May 17. Proc Natl Acad Sci U S A. 2019. PMID: 31101720 Free PMC article.
-
Deciphering clinical significance of BCL11A isoforms and protein expression roles in triple-negative breast cancer subtype.J Cancer Res Clin Oncol. 2023 Jul;149(7):3951-3963. doi: 10.1007/s00432-022-04301-w. Epub 2022 Aug 28. J Cancer Res Clin Oncol. 2023. PMID: 36030436 Free PMC article.
-
The SOX11 transcription factor is a critical regulator of basal-like breast cancer growth, invasion, and basal-like gene expression.Oncotarget. 2016 Mar 15;7(11):13106-21. doi: 10.18632/oncotarget.7437. Oncotarget. 2016. PMID: 26894864 Free PMC article.
-
Single-Cell Transcriptomic and Epigenetic Analyses of Mouse Mammary Development Starting with the Embryo.Methods Mol Biol. 2022;2471:49-82. doi: 10.1007/978-1-0716-2193-6_3. Methods Mol Biol. 2022. PMID: 35175591 Free PMC article.
References
-
- Rather J. The Genesis of Cancer: a Study in the History of Ideas. Baltimore, MD: Johns Hopkins University Press; 1978.
-
- Stephens PJ, Tarpey PS, Davies H, Van Loo P, Greenman C, Wedge DC, Nik-Zainal S, Martin S, Varela I, Bignell GR, Yates LR, Papaemmanuil E, Beare D, Butler A, Cheverton A, Gamble J, Hinton J, Jia M, Jayakumar A, Jones D, Latimer C, Lau KW, McLaren S, McBride DJ, Menzies A, Mudie L, Raine K, Rad R, Chapman MS, Teague J. et al.The landscape of cancer genes and mutational processes in breast cancer. Nature. 2012;486:400–404. - PMC - PubMed
-
- Asselin-Labat ML, Sutherland KD, Barker H, Thomas R, Shackleton M, Forrest NC, Hartley L, Robb L, Grosveld FG, van der Wees J, Lindeman GJ, Visvader JE. Gata-3 is an essential regulator of mammary-gland morphogenesis and luminal-cell differentiation. Nat Cell Biol. 2007;9:201–209. doi: 10.1038/ncb1530. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
