

Neuron-released oligomeric α-synuclein is an endogenous agonist of TLR2 for paracrine activation of microglia
- PMID: 23463005
- PMCID: PMC4089961
- DOI: 10.1038/ncomms2534
Neuron-released oligomeric α-synuclein is an endogenous agonist of TLR2 for paracrine activation of microglia
Abstract
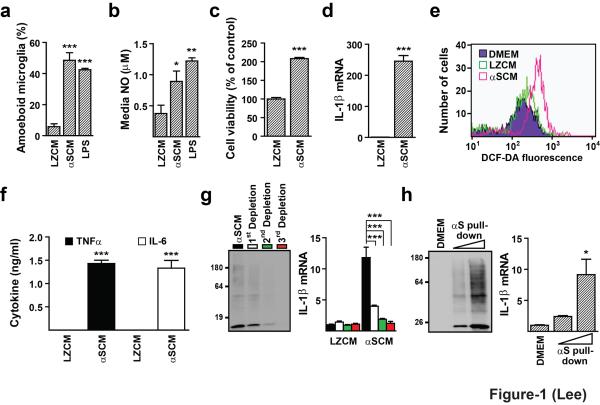
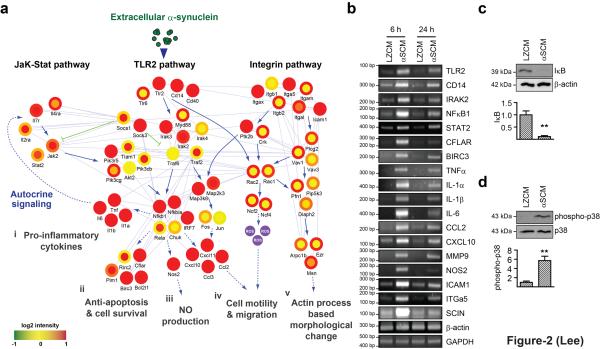
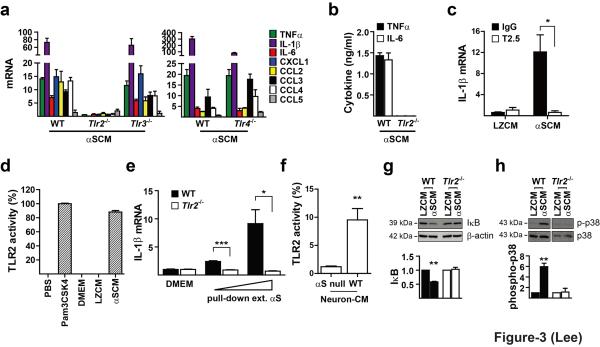
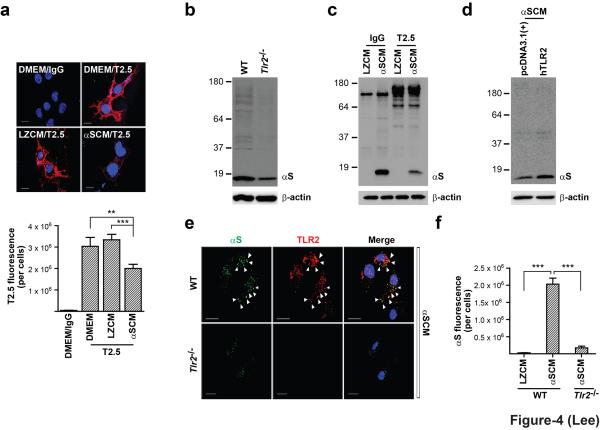
Abnormal aggregation of α-synuclein and sustained microglial activation are important contributors to the pathogenic processes of Parkinson's disease. However, the relationship between disease-associated protein aggregation and microglia-mediated neuroinflammation remains unknown. Here, using a combination of in silico, in vitro and in vivo approaches, we show that extracellular α-synuclein released from neuronal cells is an endogenous agonist for Toll-like receptor 2 (TLR2), which activates inflammatory responses in microglia. The TLR2 ligand activity of α-synuclein is conformation-sensitive; only specific types of oligomer can interact with and activate TLR2. This paracrine interaction between neuron-released oligomeric α-synuclein and TLR2 in microglia suggests that both of these proteins are novel therapeutic targets for modification of neuroinflammation in Parkinson's disease and related neurological diseases.
Figures
Similar articles
-
Reactive microglia enhance the transmission of exosomal α-synuclein via toll-like receptor 2.Brain. 2021 Aug 17;144(7):2024-2037. doi: 10.1093/brain/awab122. Brain. 2021. PMID: 33792662
-
Microglial phenotypes and toll-like receptor 2 in the substantia nigra and hippocampus of incidental Lewy body disease cases and Parkinson's disease patients.Acta Neuropathol Commun. 2014 Aug 7;2:90. doi: 10.1186/s40478-014-0090-1. Acta Neuropathol Commun. 2014. PMID: 25099483 Free PMC article.
-
β1-integrin-dependent migration of microglia in response to neuron-released α-synuclein.Exp Mol Med. 2014 Apr 18;46(4):e91. doi: 10.1038/emm.2014.6. Exp Mol Med. 2014. PMID: 24743837 Free PMC article.
-
Pathological α-synuclein exacerbates the progression of Parkinson's disease through microglial activation.Toxicol Lett. 2017 Jan 4;265:30-37. doi: 10.1016/j.toxlet.2016.11.002. Epub 2016 Nov 16. Toxicol Lett. 2017. PMID: 27865851 Review.
-
Extracellular α--synuclein-a novel and crucial factor in Lewy body diseases.Nat Rev Neurol. 2014 Feb;10(2):92-8. doi: 10.1038/nrneurol.2013.275. Epub 2014 Jan 28. Nat Rev Neurol. 2014. PMID: 24468877 Review.
Cited by
-
α-Synuclein, a chemoattractant, directs microglial migration via H2O2-dependent Lyn phosphorylation.Proc Natl Acad Sci U S A. 2015 Apr 14;112(15):E1926-35. doi: 10.1073/pnas.1417883112. Epub 2015 Mar 30. Proc Natl Acad Sci U S A. 2015. PMID: 25825709 Free PMC article.
-
Identification of a specific α-synuclein peptide (α-Syn 29-40) capable of eliciting microglial superoxide production to damage dopaminergic neurons.J Neuroinflammation. 2016 Jun 21;13(1):158. doi: 10.1186/s12974-016-0606-7. J Neuroinflammation. 2016. PMID: 27329107 Free PMC article.
-
Interrelation of oxidative stress and inflammation in neurodegenerative disease: role of TNF.Oxid Med Cell Longev. 2015;2015:610813. doi: 10.1155/2015/610813. Epub 2015 Mar 5. Oxid Med Cell Longev. 2015. PMID: 25834699 Free PMC article. Review.
-
Protein Transmission, Seeding and Degradation: Key Steps for α-Synuclein Prion-Like Propagation.Exp Neurobiol. 2014 Dec;23(4):324-36. doi: 10.5607/en.2014.23.4.324. Epub 2014 Dec 12. Exp Neurobiol. 2014. PMID: 25548532 Free PMC article. Review.
-
Glial Purinergic Signaling in Neurodegeneration.Front Neurol. 2021 May 14;12:654850. doi: 10.3389/fneur.2021.654850. eCollection 2021. Front Neurol. 2021. PMID: 34054698 Free PMC article. Review.
References
-
- Hirsch EC, Hunot S. Neuroinflammation in Parkinson's disease: a target for neuroprotection? Lancet Neurol. 2009;8:382–397. - PubMed
-
- Graeber MB, Streit WJ. Microglia: biology and pathology. Acta Neuropathol. 2010;119:89–105. - PubMed
-
- Kim C, Lee S-J. Controlling the mass action of alpha-synuclein in Parkinson's disease. J Neurochem. 2008;107:303–316. - PubMed
-
- Jang A, et al. Non-classical exocytosis of alpha-synuclein is sensitive to folding states and promoted under stress conditions. J Neurochem. 2010;113:1263–1274. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
