

Review
doi: 10.1083/jcb.201211113.
Review series: The functions and consequences of force at kinetochores
Affiliations
- PMID: 23460675
- PMCID: PMC3587826
- DOI: 10.1083/jcb.201211113
Item in Clipboard
Review
Review series: The functions and consequences of force at kinetochores
J Cell Biol.
.
Abstract
Chromosome segregation requires the generation of force at the kinetochore-the multiprotein structure that facilitates attachment of chromosomes to spindle microtubules. This force is required both to move chromosomes and to signal the formation of proper bioriented attachments. To understand the role of force in these processes, it is critical to define how force is generated at kinetochores, the contributions of this force to chromosome movement, and how the kinetochore is structured and organized to withstand and respond to force. Classical studies and recent work provide a framework to dissect the mechanisms, functions, and consequences of force at kinetochores.
Figures
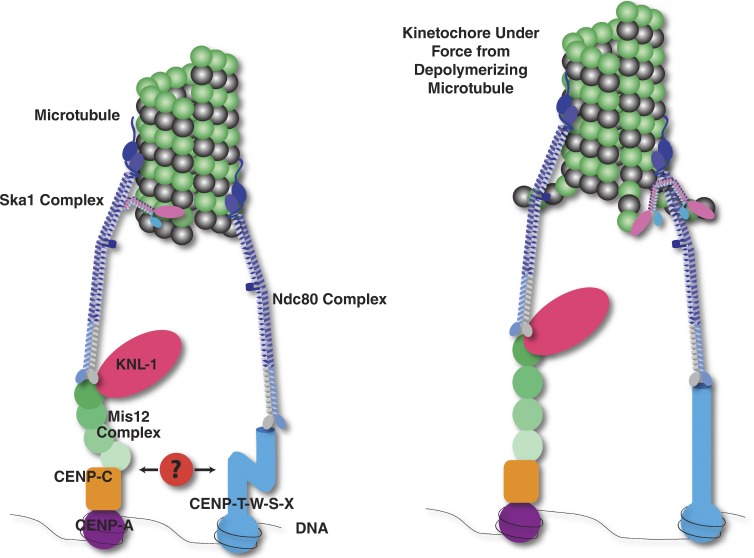
Simplified diagram of the kinetochore showing the major proteins involved in the DNA–microtubule attachment. (Left) The Ndc80 complex (dark blue) binds to microtubules and forms two separate connections to kinetochores. First, the Ndc80 complex binds to the Mis12 complex (green) and KNL-1 (magenta). The Mis12 complex in turn binds to CENP-C (orange), which binds to nucleosomes containing the histone H3 variant CENP-A (purple). Second, the Ndc80 complex binds to CENP-T (light blue). CENP-T interacts with DNA as a part of a heterotetrameric nucleosome-like CENP-T–W–S–X complex. In humans, the Ndc80 complex attachment to microtubules is enhanced by an interaction with the Ska1 complex (pink and blue; Schmidt et al., 2012). Additional components may form interactions between the two connective pathways (red). (Right) Upon microtubule depolymerization, the flexible protein components of the kinetochore may rearrange. For example, recent evidence has suggested that the N and C termini of CENP-T separate under tension (Suzuki et al., 2011) and that the subunits of the Mis12 complex redistribute (Wan et al., 2009).
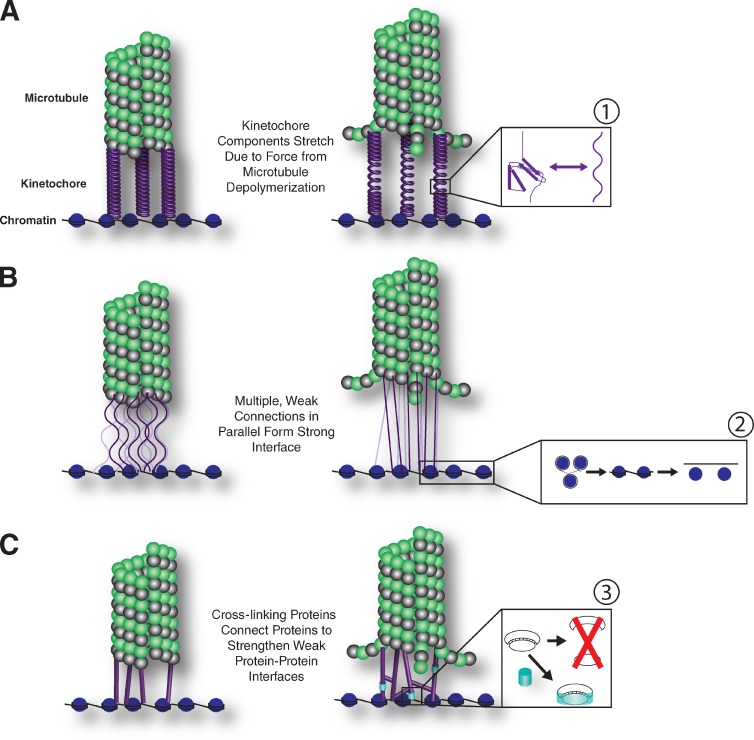
Models for force response at kinetochores at both the individual protein level and global scale. (A–C) We propose three nonexclusive models for how kinetochores respond to the application of force: kinetochore proteins with elastic properties could serve to absorb some of the force produced by depolymerizing microtubules (A), multiple weak interfaces could form parallel attachments between the depolymerizing microtubule and chromosome such that the force produced by the microtubule would be diffused across multiple connections (B), and additional kinetochore components could serve as dynamic cross-linkers to diffuse force and add interactions between pairs of proteins to strengthen the protein–protein interface (C). The kinetochore protein components themselves could have multiple responses at a molecular level including that (1) under pulling forces, the bonds holding together the tertiary and secondary structure of a protein can break, causing the protein to unfold. If reversible, this would provide elastic properties, but if permanent, could lead to loss of functional kinetochore components. (2) The force generated by kinetochores is directed toward the limited number of protein–DNA interactions formed between the kinetochore proteins and the chromosome. Some tension may be relieved as the DNA wrapped around adjacent nucleosomes is pulled. This first results in the straightening out of the compact “beads on a string” structure, but with sufficient pulling force, the nucleosomes would be removed from the DNA. (3) Protein–protein interfaces held together by noncovalent bonds can break under pulling force, but the presence of additional proteins to strengthen interactions could prevent the loss of important interfaces.
Similar articles
-
Spindle assembly checkpoint satisfaction occurs via end-on but not lateral attachments under tension.J Cell Biol. 2017 Jun 5;216(6):1533-1542. doi: 10.1083/jcb.201611104. Epub 2017 May 23. J Cell Biol. 2017. PMID: 28536121 Free PMC article.
-
Individual kinetochore-fibers locally dissipate force to maintain robust mammalian spindle structure.J Cell Biol. 2020 Aug 3;219(8):e201911090. doi: 10.1083/jcb.201911090. J Cell Biol. 2020. PMID: 32435797 Free PMC article.
-
Mechanisms of chromosome biorientation and bipolar spindle assembly analyzed by computational modeling.Elife. 2020 Feb 13;9:e48787. doi: 10.7554/eLife.48787. Elife. 2020. PMID: 32053104 Free PMC article.
-
Merotelic kinetochores in mammalian tissue cells.Philos Trans R Soc Lond B Biol Sci. 2005 Mar 29;360(1455):553-68. doi: 10.1098/rstb.2004.1610. Philos Trans R Soc Lond B Biol Sci. 2005. PMID: 15897180 Free PMC article. Review.
-
Microtubule attachment and spindle assembly checkpoint signalling at the kinetochore.Nat Rev Mol Cell Biol. 2013 Jan;14(1):25-37. doi: 10.1038/nrm3494. Nat Rev Mol Cell Biol. 2013. PMID: 23258294 Free PMC article. Review.
Cited by
-
SKAP binding to microtubules reduces friction at the kinetochore-microtubule interface and increases attachment stability under force.bioRxiv [Preprint]. 2024 Aug 8:2024.08.08.607154. doi: 10.1101/2024.08.08.607154. bioRxiv. 2024. PMID: 39149232 Free PMC article. Preprint.
-
A quantitative description of Ndc80 complex linkage to human kinetochores.Nat Commun. 2015 Sep 8;6:8161. doi: 10.1038/ncomms9161. Nat Commun. 2015. PMID: 26345214 Free PMC article.
-
Loss of Ikbkap/Elp1 in mouse oocytes causes spindle disorganization, developmental defects in preimplantation embryos and impaired female fertility.Sci Rep. 2019 Dec 11;9(1):18875. doi: 10.1038/s41598-019-55090-1. Sci Rep. 2019. PMID: 31827135 Free PMC article.
-
The kinetochore.Cold Spring Harb Perspect Biol. 2014 Jul 1;6(7):a015826. doi: 10.1101/cshperspect.a015826. Cold Spring Harb Perspect Biol. 2014. PMID: 24984773 Free PMC article. Review.
-
Ska3 Ensures Timely Mitotic Progression by Interacting Directly With Microtubules and Ska1 Microtubule Binding Domain.Sci Rep. 2016 Sep 26;6:34042. doi: 10.1038/srep34042. Sci Rep. 2016. PMID: 27667719 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
