

Crystal structure of glycoprotein C from Rift Valley fever virus
- PMID: 23319635
- PMCID: PMC3562824
- DOI: 10.1073/pnas.1217780110
Crystal structure of glycoprotein C from Rift Valley fever virus
Abstract
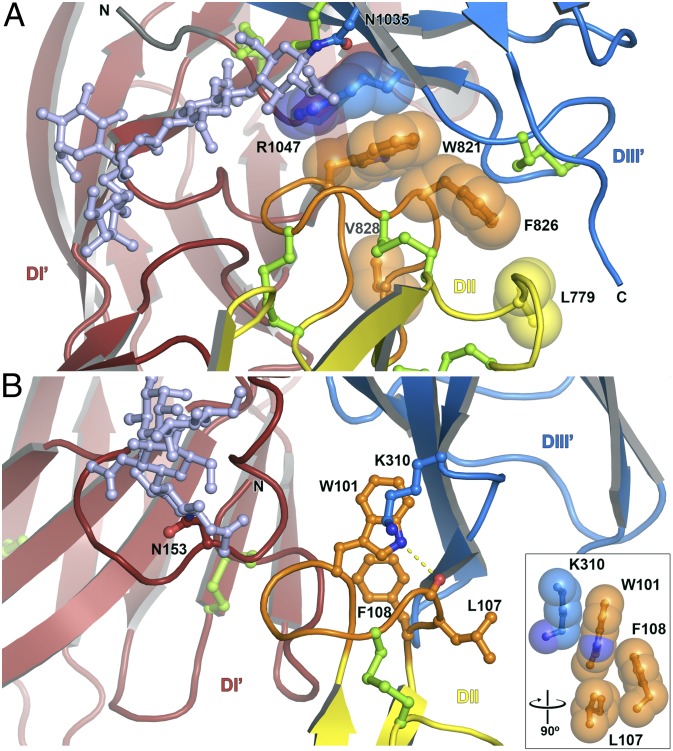
Rift Valley fever virus (RVFV), like many other Bunyaviridae family members, is an emerging human and animal pathogen. Bunyaviruses have an outer lipid envelope bearing two glycoproteins, G(N) and G(C), required for cell entry. Bunyaviruses deliver their genome into the host-cell cytoplasm by fusing their envelope with an endosomal membrane. The molecular mechanism of this key entry step is unknown. The crystal structure of RVFV G(C) reveals a class II fusion protein architecture found previously in flaviviruses and alphaviruses. The structure identifies G(C) as the effector of membrane fusion and provides a direct view of the membrane anchor that initiates fusion. A structure of nonglycosylated G(C) reveals an extended conformation that may represent a fusion intermediate. Unanticipated similarities between G(C) and flavivirus envelope proteins reveal an evolutionary link between the two virus families and provide insights into the organization of G(C) in the outer shell of RVFV.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
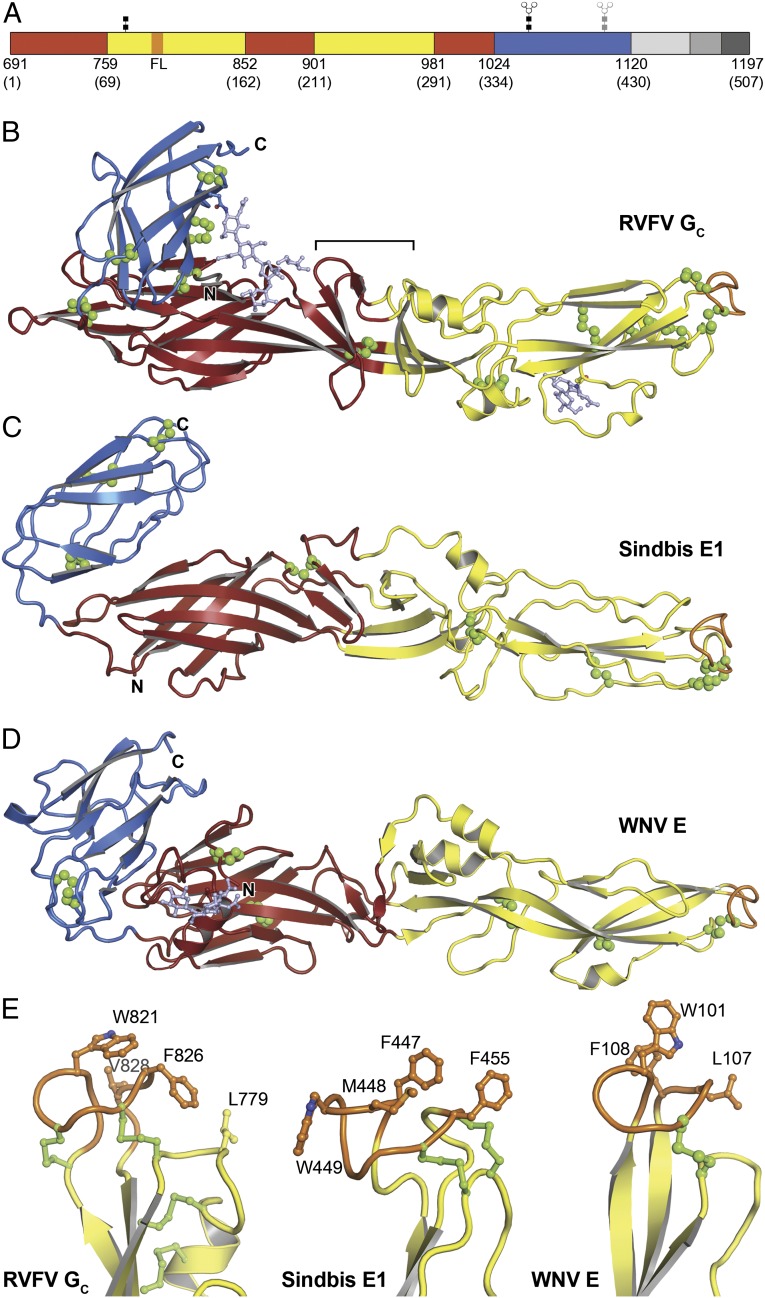
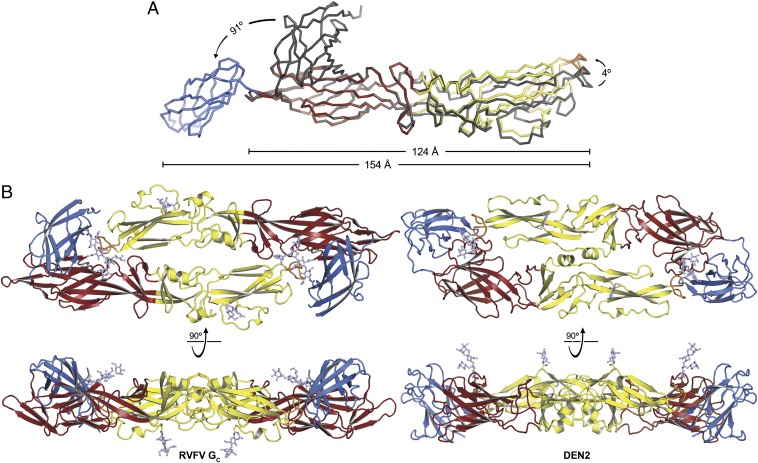
Similar articles
-
[Study on DNA immune of envelope protein gene of Rift Valley Fever Virus].Wei Sheng Wu Xue Bao. 2007 Aug;47(4):677-81. Wei Sheng Wu Xue Bao. 2007. PMID: 17944371 Chinese.
-
Structures of phlebovirus glycoprotein Gn and identification of a neutralizing antibody epitope.Proc Natl Acad Sci U S A. 2017 Sep 5;114(36):E7564-E7573. doi: 10.1073/pnas.1705176114. Epub 2017 Aug 21. Proc Natl Acad Sci U S A. 2017. PMID: 28827346 Free PMC article.
-
Structural changes of envelope proteins during alphavirus fusion.Nature. 2010 Dec 2;468(7324):705-8. doi: 10.1038/nature09546. Nature. 2010. PMID: 21124457 Free PMC article.
-
The pathogenesis of Rift Valley fever.Viruses. 2011 May;3(5):493-519. doi: 10.3390/v3050493. Viruses. 2011. PMID: 21666766 Free PMC article. Review.
-
The Role of Phlebovirus Glycoproteins in Viral Entry, Assembly and Release.Viruses. 2016 Jul 21;8(7):202. doi: 10.3390/v8070202. Viruses. 2016. PMID: 27455305 Free PMC article. Review.
Cited by
-
Rift Valley Fever Virus-Infection, Pathogenesis and Host Immune Responses.Pathogens. 2023 Sep 19;12(9):1174. doi: 10.3390/pathogens12091174. Pathogens. 2023. PMID: 37764982 Free PMC article. Review.
-
Incorporation of hepatitis C virus E1 and E2 glycoproteins: the keystones on a peculiar virion.Viruses. 2014 Mar 11;6(3):1149-87. doi: 10.3390/v6031149. Viruses. 2014. PMID: 24618856 Free PMC article. Review.
-
Shielding and activation of a viral membrane fusion protein.Nat Commun. 2018 Jan 24;9(1):349. doi: 10.1038/s41467-017-02789-2. Nat Commun. 2018. PMID: 29367607 Free PMC article.
-
Potent neutralization of Rift Valley fever virus by human monoclonal antibodies through fusion inhibition.Proc Natl Acad Sci U S A. 2021 Apr 6;118(14):e2025642118. doi: 10.1073/pnas.2025642118. Proc Natl Acad Sci U S A. 2021. PMID: 33782133 Free PMC article.
-
Crystal structure of glycoprotein E2 from bovine viral diarrhea virus.Proc Natl Acad Sci U S A. 2013 Apr 23;110(17):6805-10. doi: 10.1073/pnas.1300524110. Epub 2013 Apr 8. Proc Natl Acad Sci U S A. 2013. PMID: 23569276 Free PMC article.
References
-
- Schmaljohn CS, Nichol ST. Bunyaviridae. In: Knipe DM, Howley PM, editors. Fields Virology. Philadelphia: Lippincott Williams & Wilkins; 2007. 5th Ed, Vol 2, pp 1741–1788.
-
- Swanepoel R, Coetzer JAW. Rift Valley fever. In: Coetzer JAW, Thomson GR, Tustin RC, Kriek NPJ, editors. Infectious Diseases of Livestock: With Special Reference to Southern Africa. Cape Town: Oxford Univ Press; 1994. pp. 688–717.
-
- Gargan TP, 2nd, Clark GG, Dohm DJ, Turell MJ, Bailey CL. Vector potential of selected North American mosquito species for Rift Valley fever virus. Am J Trop Med Hyg. 1988;38(2):440–446. - PubMed
-
- Enserink M. Entomology. A mosquito goes global. Science. 2008;320(5878):864–866. - PubMed
-
- Stone R. Infectious diseases. Rival teams identify a virus behind deaths in central China. Science. 2010;330(6000):20–21. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
