

Cellular and molecular mechanisms of muscle atrophy
- PMID: 23268536
- PMCID: PMC3529336
- DOI: 10.1242/dmm.010389
Cellular and molecular mechanisms of muscle atrophy
Abstract
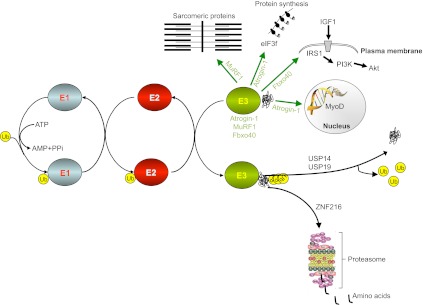
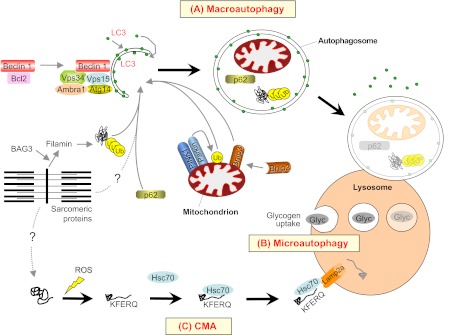
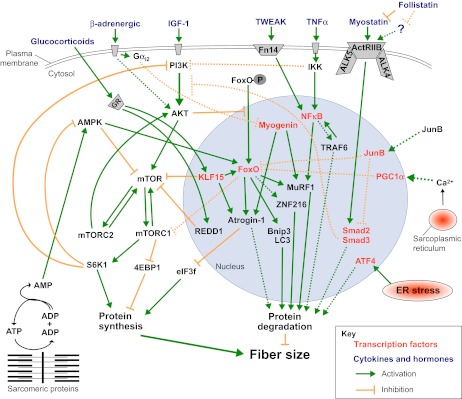
Skeletal muscle is a plastic organ that is maintained by multiple pathways regulating cell and protein turnover. During muscle atrophy, proteolytic systems are activated, and contractile proteins and organelles are removed, resulting in the shrinkage of muscle fibers. Excessive loss of muscle mass is associated with poor prognosis in several diseases, including myopathies and muscular dystrophies, as well as in systemic disorders such as cancer, diabetes, sepsis and heart failure. Muscle loss also occurs during aging. In this paper, we review the key mechanisms that regulate the turnover of contractile proteins and organelles in muscle tissue, and discuss how impairments in these mechanisms can contribute to muscle atrophy. We also discuss how protein synthesis and degradation are coordinately regulated by signaling pathways that are influenced by mechanical stress, physical activity, and the availability of nutrients and growth factors. Understanding how these pathways regulate muscle mass will provide new therapeutic targets for the prevention and treatment of muscle atrophy in metabolic and neuromuscular diseases.
Figures
Similar articles
-
Protein breakdown in muscle wasting: role of autophagy-lysosome and ubiquitin-proteasome.Int J Biochem Cell Biol. 2013 Oct;45(10):2121-9. doi: 10.1016/j.biocel.2013.04.023. Epub 2013 May 7. Int J Biochem Cell Biol. 2013. PMID: 23665154 Free PMC article. Review.
-
Misregulation of autophagy and protein degradation systems in myopathies and muscular dystrophies.J Cell Sci. 2013 Dec 1;126(Pt 23):5325-33. doi: 10.1242/jcs.114041. J Cell Sci. 2013. PMID: 24293330 Review.
-
FoxO3 coordinately activates protein degradation by the autophagic/lysosomal and proteasomal pathways in atrophying muscle cells.Cell Metab. 2007 Dec;6(6):472-83. doi: 10.1016/j.cmet.2007.11.004. Cell Metab. 2007. PMID: 18054316
-
Regulation of autophagy and the ubiquitin-proteasome system by the FoxO transcriptional network during muscle atrophy.Nat Commun. 2015 Apr 10;6:6670. doi: 10.1038/ncomms7670. Nat Commun. 2015. PMID: 25858807 Free PMC article.
-
P38α MAPK coordinates the activities of several metabolic pathways that together induce atrophy of denervated muscles.FEBS J. 2020 Jan;287(1):73-93. doi: 10.1111/febs.15070. Epub 2019 Oct 22. FEBS J. 2020. PMID: 31545558
Cited by
-
Insights into the key roles of proteoglycans in breast cancer biology and translational medicine.Biochim Biophys Acta. 2015 Apr;1855(2):276-300. doi: 10.1016/j.bbcan.2015.03.006. Epub 2015 Mar 28. Biochim Biophys Acta. 2015. PMID: 25829250 Free PMC article. Review.
-
Vitamin D3 Exerts Beneficial Effects on C2C12 Myotubes through Activation of the Vitamin D Receptor (VDR)/Sirtuins (SIRT)1/3 Axis.Nutrients. 2023 Nov 7;15(22):4714. doi: 10.3390/nu15224714. Nutrients. 2023. PMID: 38004107 Free PMC article.
-
The SH3 and cysteine-rich domain 3 (Stac3) gene is important to growth, fiber composition, and calcium release from the sarcoplasmic reticulum in postnatal skeletal muscle.Skelet Muscle. 2016 Apr 11;6:17. doi: 10.1186/s13395-016-0088-4. eCollection 2016. Skelet Muscle. 2016. PMID: 27073615 Free PMC article.
-
Inositol Hexakisphosphate Kinase 3 Regulates Metabolism and Lifespan in Mice.Sci Rep. 2016 Aug 31;6:32072. doi: 10.1038/srep32072. Sci Rep. 2016. PMID: 27577108 Free PMC article.
-
Development of a local controlled release system for therapeutic proteins in the treatment of skeletal muscle injuries and diseases.Cell Death Dis. 2024 Jul 2;15(7):470. doi: 10.1038/s41419-024-06645-2. Cell Death Dis. 2024. PMID: 38956034 Free PMC article.
References
-
- Allen D. L., Unterman T. G. (2007). Regulation of myostatin expression and myoblast differentiation by FoxO and SMAD transcription factors. Am. J. Physiol. Cell. Physiol. 292, C188–C199 - PubMed
-
- Amirouche A., Durieux A. C., Banzet S., Koulmann N., Bonnefoy R., Mouret C., Bigard X., Peinnequin A., Freyssenet D. (2009). Down-regulation of Akt/mammalian target of rapamycin signaling pathway in response to myostatin overexpression in skeletal muscle. Endocrinology 150, 286–294 - PubMed
-
- Arndt V., Dick N., Tawo R., Dreiseidler M., Wenzel D., Hesse M., Fürst D. O., Saftig P., Saint R., Fleischmann B. K., et al. (2010). Chaperone-assisted selective autophagy is essential for muscle maintenance. Curr. Biol. 20, 143–148 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
