

A ribosome-specialized translation initiation pathway is required for cap-dependent translation of vesicular stomatitis virus mRNAs
- PMID: 23169626
- PMCID: PMC3538191
- DOI: 10.1073/pnas.1216454109
A ribosome-specialized translation initiation pathway is required for cap-dependent translation of vesicular stomatitis virus mRNAs
Abstract
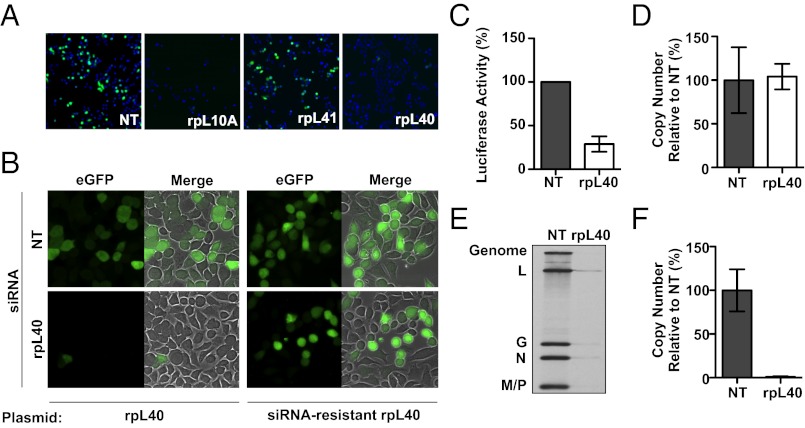
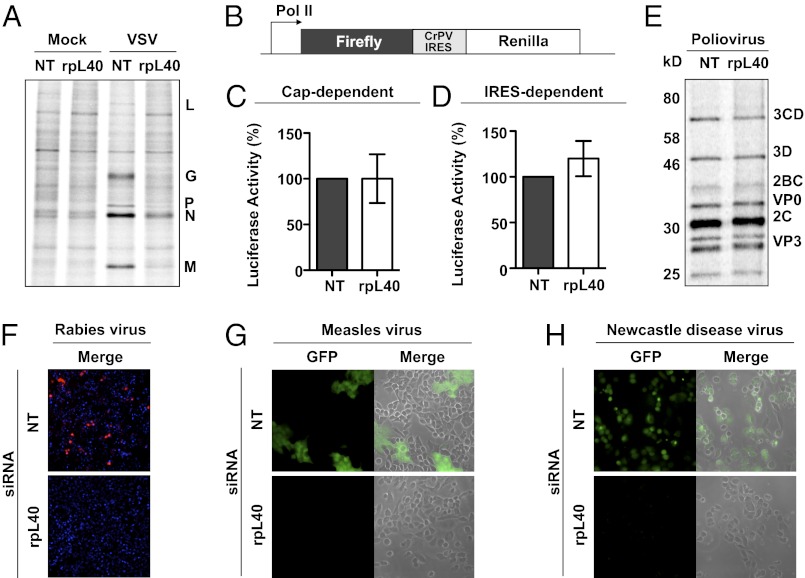
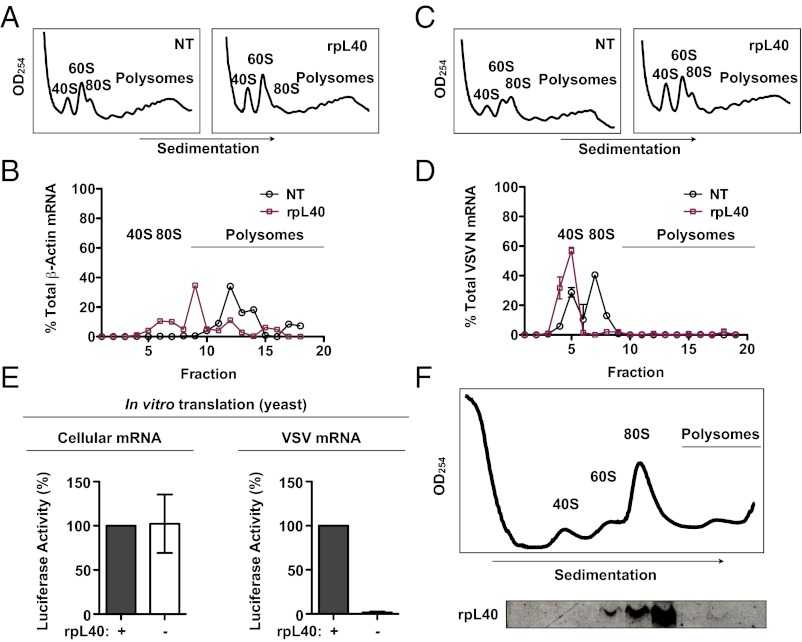
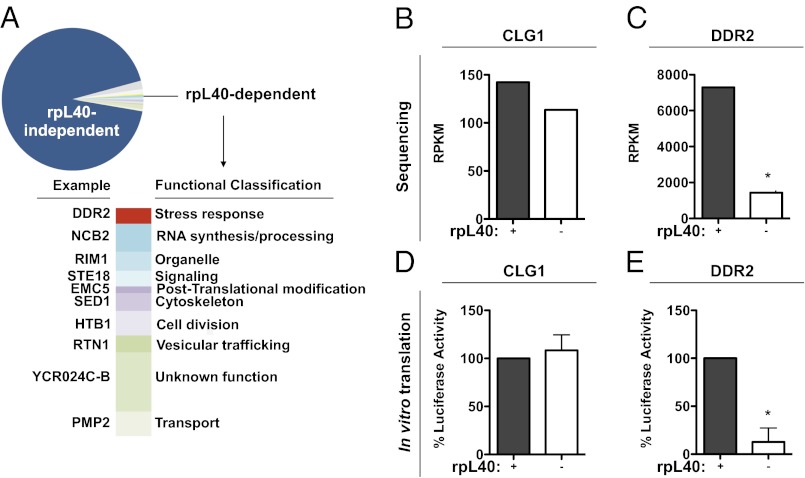
Initiation is the primary target of translational control for all organisms. Regulation of eukaryotic translation is traditionally thought to occur through initiation factors and RNA structures. Here, we characterize a transcript-specific translation initiation mechanism that is mediated by the ribosome. By studying vesicular stomatitis virus (VSV), we identify the large ribosomal subunit protein rpL40 as requisite for VSV cap-dependent translation but not bulk cellular or internal ribosome entry site-driven translation. This requirement is conserved among members of the order Mononegavirales, including measles virus and rabies virus. Polysome analyses and in vitro reconstitution of initiation demonstrate that rpL40 is required for 80S formation on VSV mRNAs through a cis-regulatory element. Using deep sequencing, we further uncover a subset of cellular transcripts that are selectively sensitive to rpL40 depletion, suggesting VSV may have usurped an endogenous translation pathway. Together, these findings demonstrate that the ribosome acts as a translational regulator outside of its catalytic role during protein synthesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Ribosomes take control.Proc Natl Acad Sci U S A. 2013 Jan 2;110(1):9-10. doi: 10.1073/pnas.1218764110. Epub 2012 Dec 14. Proc Natl Acad Sci U S A. 2013. PMID: 23243144 Free PMC article. No abstract available.
Similar articles
-
Ribosomes take control.Proc Natl Acad Sci U S A. 2013 Jan 2;110(1):9-10. doi: 10.1073/pnas.1218764110. Epub 2012 Dec 14. Proc Natl Acad Sci U S A. 2013. PMID: 23243144 Free PMC article. No abstract available.
-
Ribosomal Protein L13 Promotes IRES-Driven Translation of Foot-and-Mouth Disease Virus in a Helicase DDX3-Dependent Manner.J Virol. 2020 Jan 6;94(2):e01679-19. doi: 10.1128/JVI.01679-19. Print 2020 Jan 6. J Virol. 2020. PMID: 31619563 Free PMC article.
-
Characterization of the Interaction between hantavirus nucleocapsid protein (N) and ribosomal protein S19 (RPS19).J Biol Chem. 2011 Apr 1;286(13):11814-24. doi: 10.1074/jbc.M110.210179. Epub 2011 Feb 4. J Biol Chem. 2011. PMID: 21296889 Free PMC article.
-
mRNAs that specifically interact with eukaryotic ribosomal subunits.Biochimie. 2015 Jul;114:48-57. doi: 10.1016/j.biochi.2014.12.008. Epub 2014 Dec 18. Biochimie. 2015. PMID: 25530261 Review.
-
Regulation of translation initiation by amino acids in eukaryotic cells.Prog Mol Subcell Biol. 2001;26:155-84. doi: 10.1007/978-3-642-56688-2_6. Prog Mol Subcell Biol. 2001. PMID: 11575165 Review.
Cited by
-
Ribosome Structural Changes Dynamically Affect Ribosome Function.Int J Mol Sci. 2024 Oct 17;25(20):11186. doi: 10.3390/ijms252011186. Int J Mol Sci. 2024. PMID: 39456968 Free PMC article. Review.
-
Newcastle Disease virus infection activates PI3K/Akt/mTOR and p38 MAPK/Mnk1 pathways to benefit viral mRNA translation via interaction of the viral NP protein and host eIF4E.PLoS Pathog. 2020 Jun 30;16(6):e1008610. doi: 10.1371/journal.ppat.1008610. eCollection 2020 Jun. PLoS Pathog. 2020. PMID: 32603377 Free PMC article.
-
Identification of Changing Ribosome Protein Compositions using Mass Spectrometry.Proteomics. 2018 Oct;18(20):e1800217. doi: 10.1002/pmic.201800217. Proteomics. 2018. PMID: 30211483 Free PMC article.
-
A broad-spectrum antiviral molecule, QL47, selectively inhibits eukaryotic translation.J Biol Chem. 2020 Feb 7;295(6):1694-1703. doi: 10.1074/jbc.RA119.011132. Epub 2019 Dec 30. J Biol Chem. 2020. PMID: 31914414 Free PMC article.
-
Genetics. Mysterious ribosomopathies.Science. 2013 Aug 23;341(6148):849-50. doi: 10.1126/science.1244156. Science. 2013. PMID: 23970686 Free PMC article.
References
-
- Gingras AC, Raught B, Sonenberg N. eIF4 initiation factors: Effectors of mRNA recruitment to ribosomes and regulators of translation. Annu Rev Biochem. 1999;68:913–963. - PubMed
-
- Pelletier J, Sonenberg N. Internal initiation of translation of eukaryotic mRNA directed by a sequence derived from poliovirus RNA. Nature. 1988;334(6180):320–325. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
