

ER stress activates NF-κB by integrating functions of basal IKK activity, IRE1 and PERK
- PMID: 23110043
- PMCID: PMC3482226
- DOI: 10.1371/journal.pone.0045078
ER stress activates NF-κB by integrating functions of basal IKK activity, IRE1 and PERK
Abstract
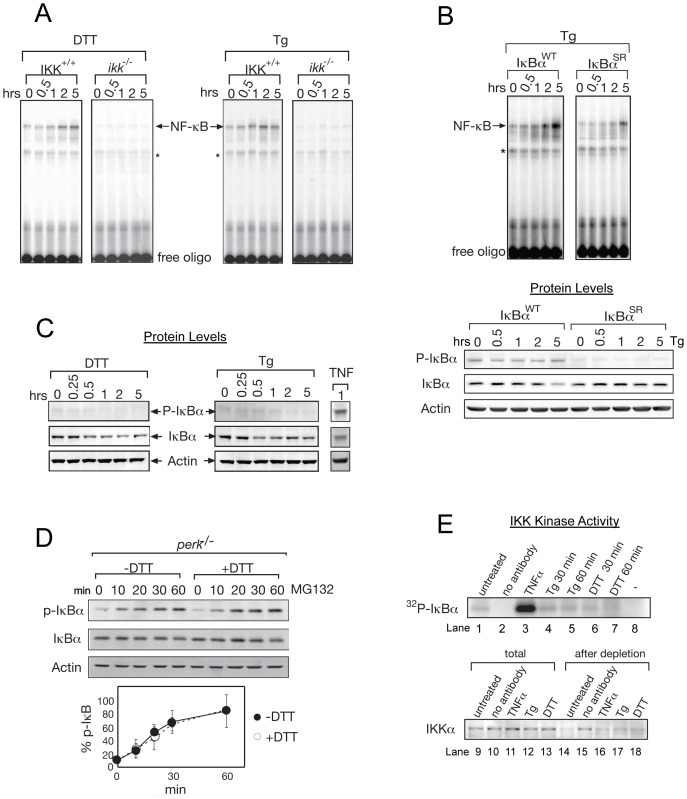
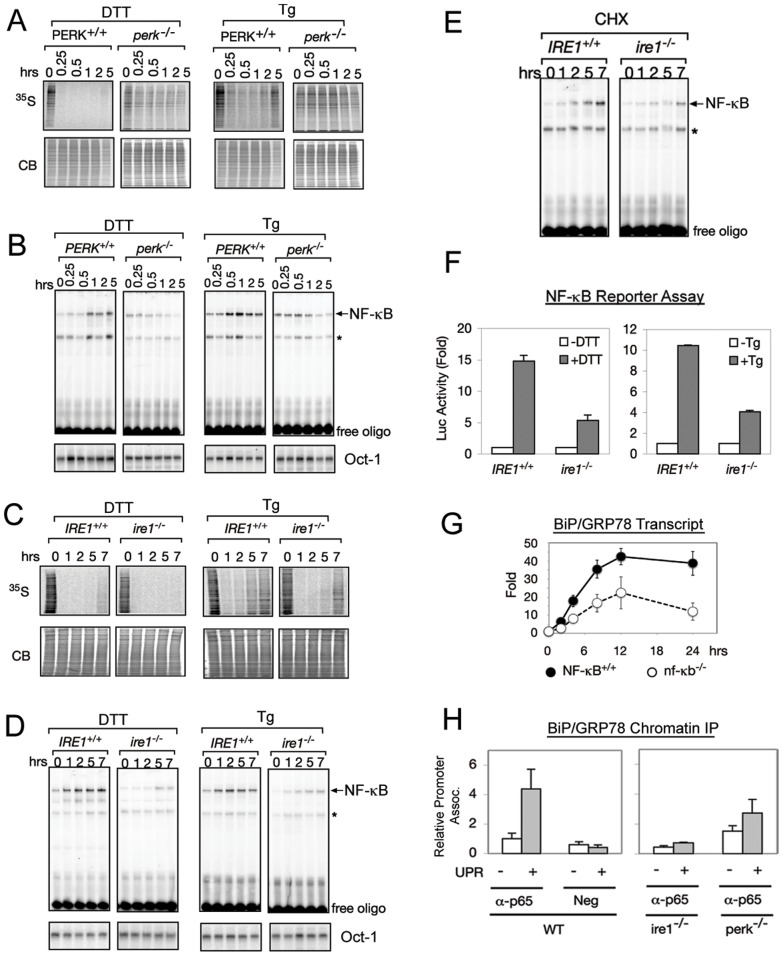
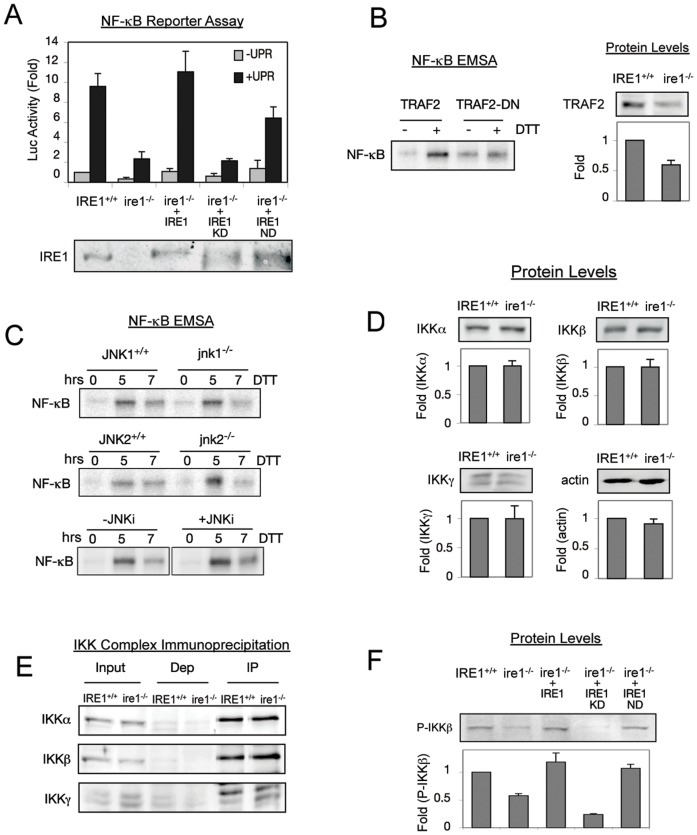
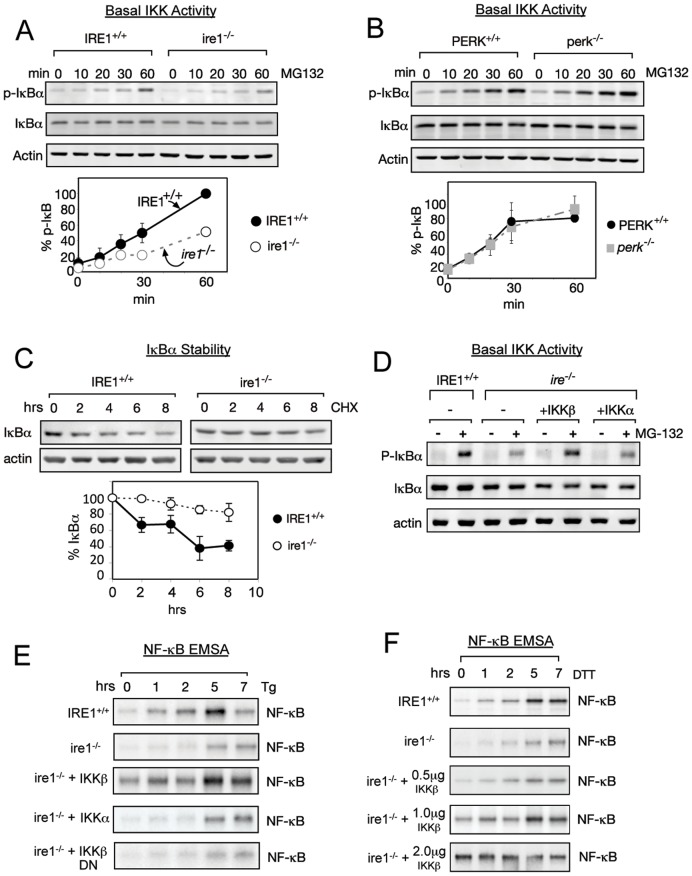
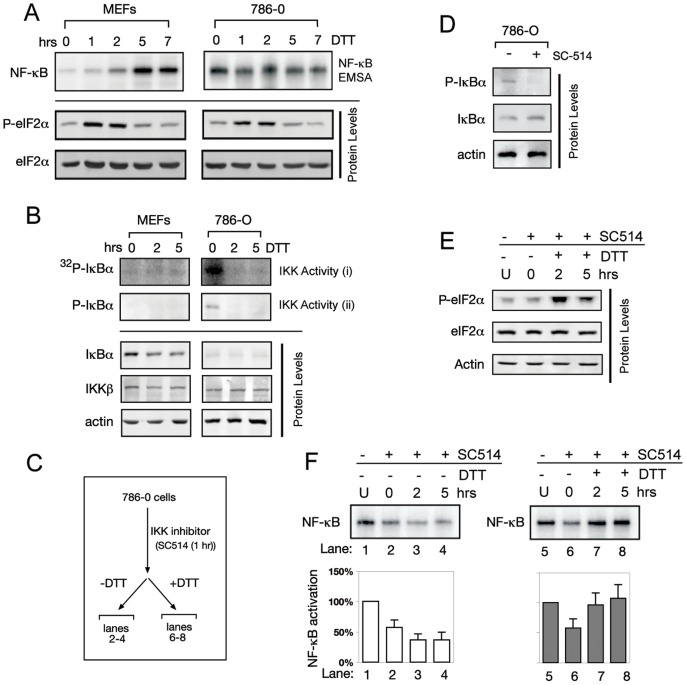
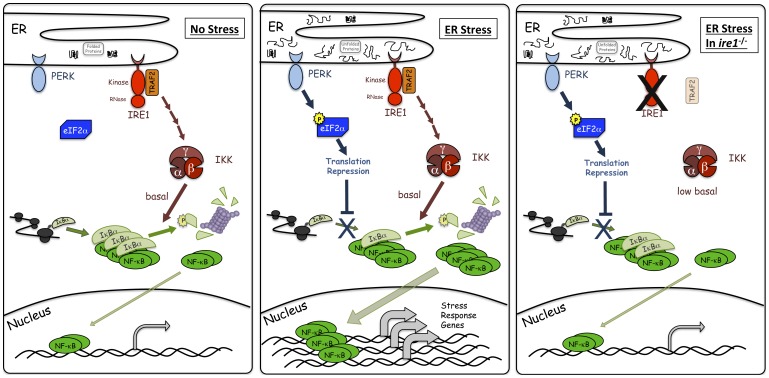
NF-κB, a transcription factor, becomes activated during the Unfolded Protein Response (UPR), an endoplasmic reticulum (ER) stress response pathway. NF-κB is normally held inactive by its inhibitor, IκBα. Multiple cellular pathways activate IKK (IκBα Kinase) which phosphorylate IκBα leading to its degradation and NF-κB activation. Here, we find that IKK is required for maximum activation of NF-κB in response to ER stress. However, unlike canonical NFκB activation, IKK activity does not increase during ER stress, but rather the level of basal IKK activity is critical for determining the extent of NF-κB activation. Furthermore, a key UPR initiator, IRE1, acts to maintain IKK basal activity through IRE1's kinase, but not RNase, activity. Inputs from IRE1 and IKK, in combination with translation repression by PERK, another UPR initiator, lead to maximal NF-κB activation during the UPR. These interdependencies have a significant impact in cancer cells with elevated IKK/NF-κB activity such as renal cell carcinoma cells (786-0). Inhibition of IKK by an IKK inhibitor, which significantly decreases NF-κB activity, is overridden by UPR induction, arguing for the importance of considering UPR activation in cancer treatment.
Conflict of interest statement
Figures
Similar articles
-
HCV induces transforming growth factor β1 through activation of endoplasmic reticulum stress and the unfolded protein response.Sci Rep. 2016 Mar 1;6:22487. doi: 10.1038/srep22487. Sci Rep. 2016. PMID: 26927933 Free PMC article.
-
Autocrine tumor necrosis factor alpha links endoplasmic reticulum stress to the membrane death receptor pathway through IRE1alpha-mediated NF-kappaB activation and down-regulation of TRAF2 expression.Mol Cell Biol. 2006 Apr;26(8):3071-84. doi: 10.1128/MCB.26.8.3071-3084.2006. Mol Cell Biol. 2006. PMID: 16581782 Free PMC article.
-
Coordination between Two Branches of the Unfolded Protein Response Determines Apoptotic Cell Fate.Mol Cell. 2018 Aug 16;71(4):629-636.e5. doi: 10.1016/j.molcel.2018.06.038. Mol Cell. 2018. PMID: 30118681
-
Endoplasmic reticulum stress sensing in the unfolded protein response.Cold Spring Harb Perspect Biol. 2013 Mar 1;5(3):a013169. doi: 10.1101/cshperspect.a013169. Cold Spring Harb Perspect Biol. 2013. PMID: 23388626 Free PMC article. Review.
-
Sensing endoplasmic reticulum stress.Adv Exp Med Biol. 2012;738:153-68. doi: 10.1007/978-1-4614-1680-7_10. Adv Exp Med Biol. 2012. PMID: 22399379 Review.
Cited by
-
Novel insight on IRE1 in the regulation of chondrocyte dedifferentiation through ER stress independent pathway.J Physiol Biochem. 2024 May;80(2):337-347. doi: 10.1007/s13105-024-01008-z. Epub 2024 Feb 10. J Physiol Biochem. 2024. PMID: 38336929
-
Aberrant lipid metabolism in anaplastic thyroid carcinoma reveals stearoyl CoA desaturase 1 as a novel therapeutic target.J Clin Endocrinol Metab. 2015 May;100(5):E697-709. doi: 10.1210/jc.2014-2764. Epub 2015 Feb 12. J Clin Endocrinol Metab. 2015. PMID: 25675381 Free PMC article.
-
MicroRNA-1291-mediated silencing of IRE1α enhances Glypican-3 expression.RNA. 2013 Jun;19(6):778-88. doi: 10.1261/rna.036483.112. Epub 2013 Apr 18. RNA. 2013. PMID: 23598528 Free PMC article.
-
Nuclear factor-kappa B-dependent X-box binding protein 1 signalling promotes the proliferation of nucleus pulposus cells under tumour necrosis factor alpha stimulation.Cell Prolif. 2019 Mar;52(2):e12542. doi: 10.1111/cpr.12542. Epub 2018 Nov 14. Cell Prolif. 2019. PMID: 30430692 Free PMC article.
-
Neuroinflammation and ER-stress are key mechanisms of acute bilirubin toxicity and hearing loss in a mouse model.PLoS One. 2018 Aug 14;13(8):e0201022. doi: 10.1371/journal.pone.0201022. eCollection 2018. PLoS One. 2018. PMID: 30106954 Free PMC article.
References
-
- Mori K (2000) Tripartite management of unfolded proteins in the endoplasmic reticulum. Cell 101: 451–454. - PubMed
-
- Ron D, Walter P (2007) Signal integration in the endoplasmic reticulum unfolded protein response. Nat Rev Mol Cell Biol 8: 519–529. - PubMed
-
- Romero-Ramirez L, Cao H, Nelson D, Hammond E, Lee AH, et al. (2004) XBP1 is essential for survival under hypoxic conditions and is required for tumor growth. Cancer Res 64: 5943–5947. - PubMed
