

Arabidopsis proline-rich protein important for development and abiotic stress tolerance is involved in microRNA biogenesis
- PMID: 23071326
- PMCID: PMC3497810
- DOI: 10.1073/pnas.1216199109
Arabidopsis proline-rich protein important for development and abiotic stress tolerance is involved in microRNA biogenesis
Abstract
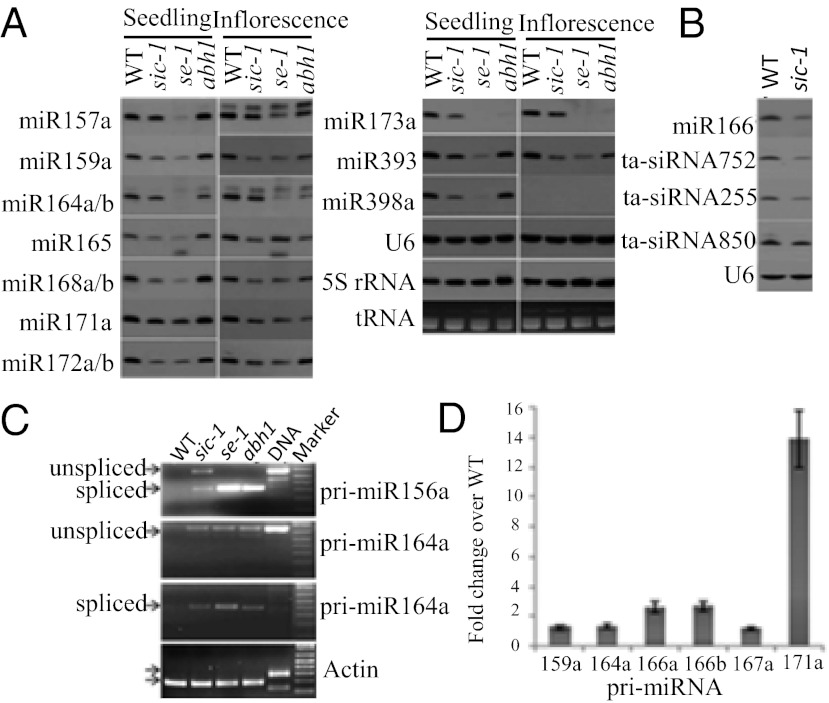
MicroRNAs (miRNAs) are important for plant development and stress responses. However, factors regulating miRNA metabolism are not completely understood. SICKLE (SIC), a proline-rich protein critical for development and abiotic stress tolerance of Arabidopsis, was identified in this study. Loss-of-function sic-1 mutant plants exhibited a serrated, sickle-like leaf margin, reduced height, delayed flowering, and abnormal inflorescence phyllotaxy, which are common characteristics of mutants involved in miRNA biogenesis. The sic-1 mutant plants accumulated lower levels of a subset of miRNAs and transacting siRNAs but higher levels of corresponding primary miRNAs than the WT. The SIC protein colocalizes with the miRNA biogenesis component HYL1 in distinct subnuclear bodies. sic-1 mutant plants also accumulated higher levels of introns from hundreds of loci. In addition, sic-1 mutant plants are hypersensitive to chilling and salt stresses. These results suggest that SIC is a unique factor required for the biogenesis of some miRNAs and degradation of some spliced introns and important for plant development and abiotic stress responses.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Similar articles
-
Strontium stress disrupts miRNA biogenesis by reducing HYL1 protein levels in Arabidopsis.Ecotoxicol Environ Saf. 2020 Nov;204:111056. doi: 10.1016/j.ecoenv.2020.111056. Epub 2020 Aug 4. Ecotoxicol Environ Saf. 2020. PMID: 32763566
-
PCA22 acts as a suppressor of atrzf1 to mediate proline accumulation in response to abiotic stress in Arabidopsis.J Exp Bot. 2017 Mar 1;68(7):1797-1809. doi: 10.1093/jxb/erx069. J Exp Bot. 2017. PMID: 28369480 Free PMC article.
-
Gene structures and processing of Arabidopsis thaliana HYL1-dependent pri-miRNAs.Nucleic Acids Res. 2009 May;37(9):3083-93. doi: 10.1093/nar/gkp189. Epub 2009 Mar 20. Nucleic Acids Res. 2009. PMID: 19304749 Free PMC article.
-
Intron Lariat RNA Inhibits MicroRNA Biogenesis by Sequestering the Dicing Complex in Arabidopsis.PLoS Genet. 2016 Nov 21;12(11):e1006422. doi: 10.1371/journal.pgen.1006422. eCollection 2016 Nov. PLoS Genet. 2016. PMID: 27870853 Free PMC article.
-
KETCH1 imports HYL1 to nucleus for miRNA biogenesis in Arabidopsis.Proc Natl Acad Sci U S A. 2017 Apr 11;114(15):4011-4016. doi: 10.1073/pnas.1619755114. Epub 2017 Mar 27. Proc Natl Acad Sci U S A. 2017. PMID: 28348234 Free PMC article.
Cited by
-
Identification and analysis of proline-rich proteins and hybrid proline-rich proteins super family genes from Sorghum bicolor and their expression patterns to abiotic stress and zinc stimuli.Front Plant Sci. 2022 Sep 26;13:952732. doi: 10.3389/fpls.2022.952732. eCollection 2022. Front Plant Sci. 2022. PMID: 36226297 Free PMC article.
-
microRNA biogenesis and turnover in plants.Cold Spring Harb Symp Quant Biol. 2012;77:183-94. doi: 10.1101/sqb.2013.77.014530. Epub 2013 Feb 25. Cold Spring Harb Symp Quant Biol. 2012. PMID: 23439913 Free PMC article. Review.
-
DAWDLE Interacts with DICER-LIKE Proteins to Mediate Small RNA Biogenesis.Plant Physiol. 2018 Jul;177(3):1142-1151. doi: 10.1104/pp.18.00354. Epub 2018 May 21. Plant Physiol. 2018. PMID: 29784765 Free PMC article.
-
STV1, a ribosomal protein, binds primary microRNA transcripts to promote their interaction with the processing complex in Arabidopsis.Proc Natl Acad Sci U S A. 2017 Feb 7;114(6):1424-1429. doi: 10.1073/pnas.1613069114. Epub 2017 Jan 23. Proc Natl Acad Sci U S A. 2017. PMID: 28115696 Free PMC article.
-
Role of proline in cell wall synthesis and plant development and its implications in plant ontogeny.Front Plant Sci. 2015 Jul 20;6:544. doi: 10.3389/fpls.2015.00544. eCollection 2015. Front Plant Sci. 2015. PMID: 26257754 Free PMC article. Review.
References
-
- Jones-Rhoades MW, Bartel DP. Computational identification of plant microRNAs and their targets, including a stress-induced miRNA. Mol Cell. 2004;14(6):787–799. - PubMed
-
- Sunkar R, Chinnusamy V, Zhu J, Zhu JK. Small RNAs as big players in plant abiotic stress responses and nutrient deprivation. Trends Plant Sci. 2007;12(7):301–309. - PubMed
-
- Voinnet O. Origin, biogenesis, and activity of plant microRNAs. Cell. 2009;136(4):669–687. - PubMed
-
- Rubio-Somoza I, Cuperus JT, Weigel D, Carrington JC. Regulation and functional specialization of small RNA-target nodes during plant development. Curr Opin Plant Biol. 2009;12(5):622–627. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
